

Acquisition of oral microbiota is driven by environment, not host genetics
- PMID: 33622378
- PMCID: PMC7903647
- DOI: 10.1186/s40168-020-00986-8
Acquisition of oral microbiota is driven by environment, not host genetics
Abstract
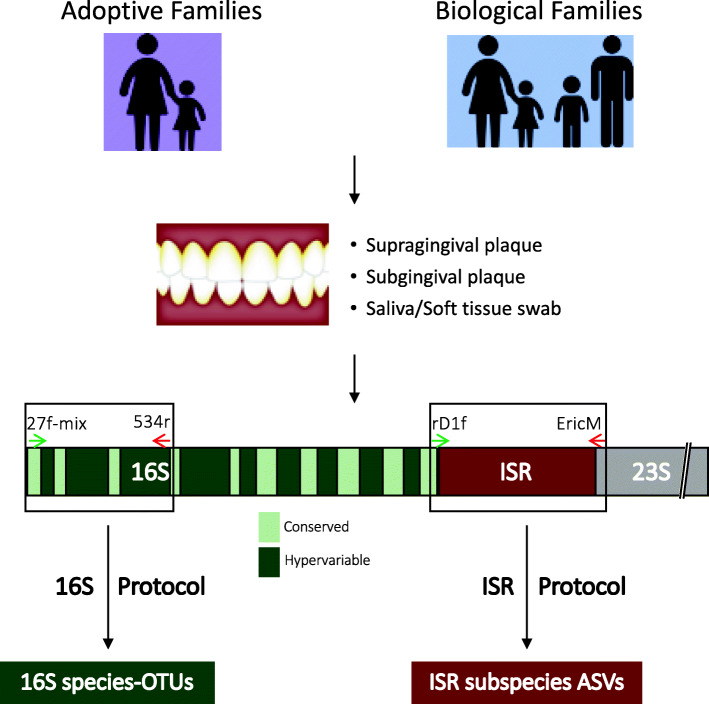
Background: The oral microbiota is acquired very early, but the factors shaping its acquisition are not well understood. Previous studies comparing monozygotic (MZ) and dizygotic (DZ) twins have suggested that host genetics plays a role. However, all twins share an equal portion of their parent's genome, so this model is not informative for studying parent-to-child transmission. We used a novel study design that allowed us to directly examine the genetics of transmission by comparing the oral microbiota of biological versus adoptive mother-child dyads.
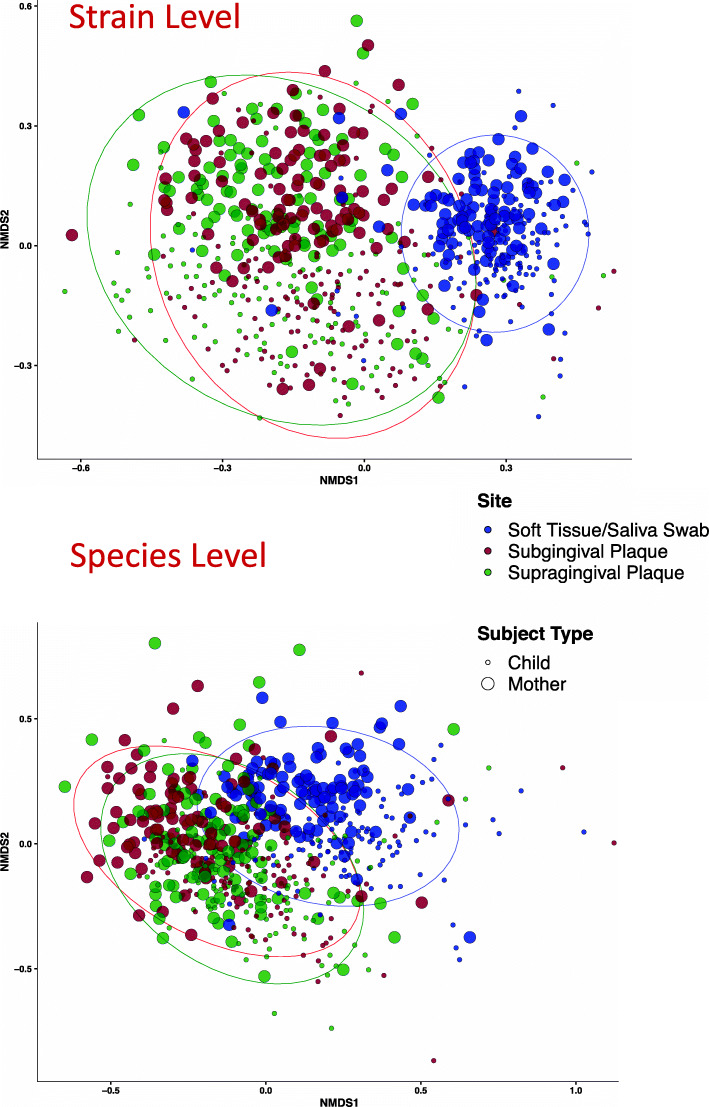
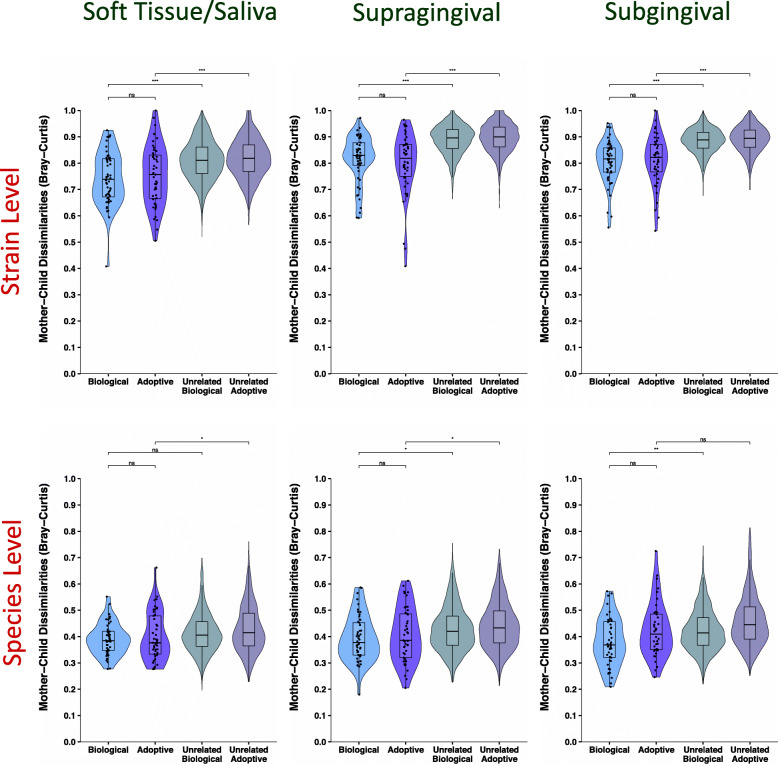
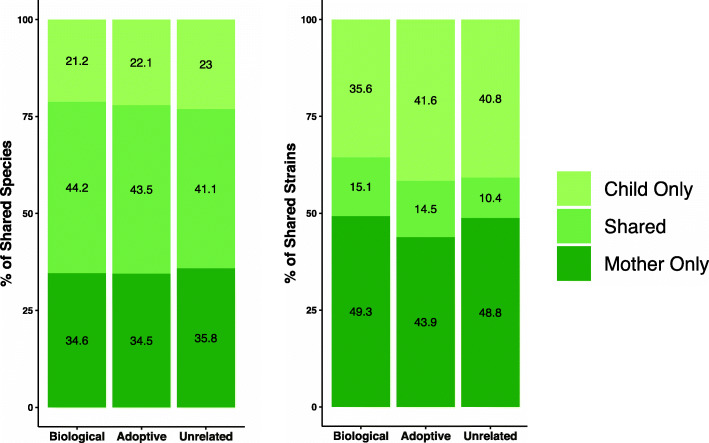
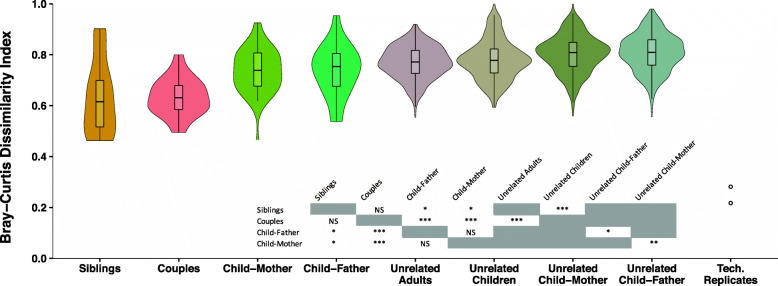
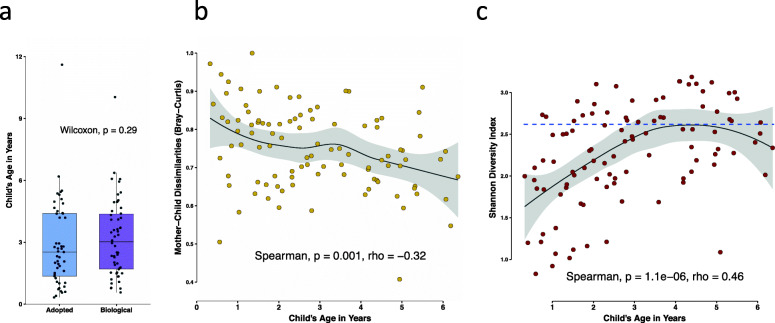
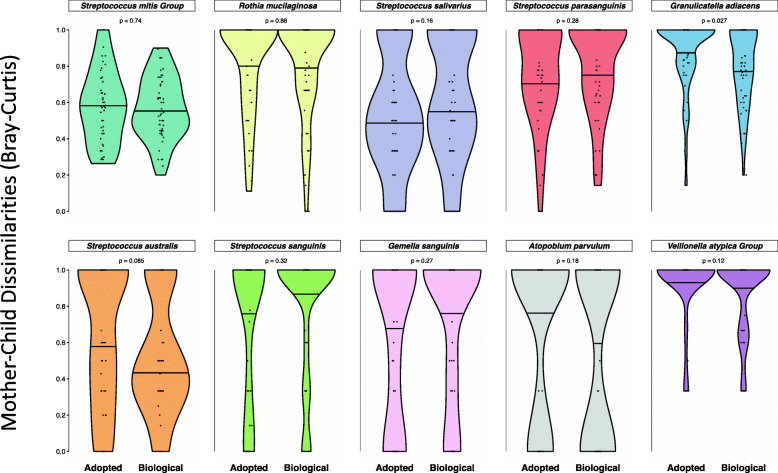
Results: No difference was observed in how closely oral bacterial community profiles matched for adoptive versus biological mother-child pairs, indicating little if any effect of host genetics on the fidelity of transmission. Both adopted and biologic children more closely resembled their own mother as compared to unrelated women, supporting the role of contact and environment. Mother-child strain similarity increased with the age of the child, ruling out early effects of host genetic influence that are lost over time. No effect on the fidelity of mother-child strain sharing from vaginal birth or breast feeding was seen. Analysis of extended families showed that fathers and mothers were equally similar to their children, and that cohabitating couples showed even greater strain similarity than mother-child pairs. These findings support the role of contact and shared environment, and age, but not genetics, as determinants of microbial transmission, and were consistent at both species and strain level resolutions, and across multiple oral habitats. In addition, analysis of individual species all showed similar results.
Conclusions: The host is clearly active in shaping the composition of the oral microbiome, since only a few of the many bacterial species in the larger environment are capable of colonizing the human oral cavity. Our findings suggest that these host mechanisms are universally shared among humans, since no effect of genetic relatedness on fidelity of microbial transmission could be detected. Instead our findings point towards contact and shared environment being the driving factors of microbial transmission, with a unique combination of these factors ultimately shaping the highly personalized human oral microbiome. Video abstract.
Keywords: Acquisition; Genetics; Human; Oral microbiome; Strain level; Transmission.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
