

Parameters Affecting Continuous In Vitro Culture of Treponema pallidum Strains
- PMID: 33622721
- PMCID: PMC8545124
- DOI: 10.1128/mBio.03536-20
Parameters Affecting Continuous In Vitro Culture of Treponema pallidum Strains
Abstract
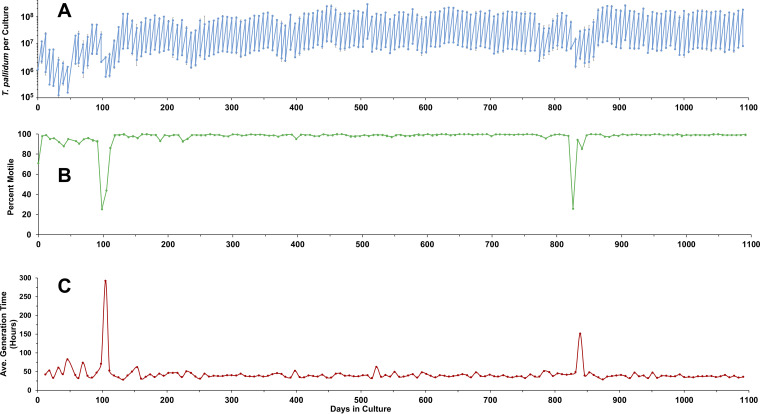
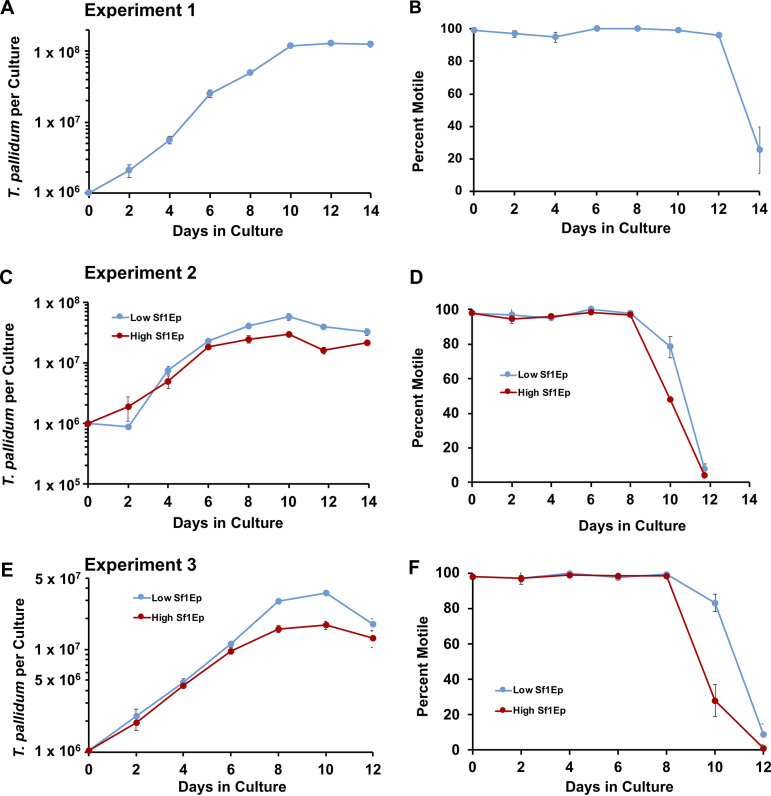
The bacterium that causes syphilis, Treponema pallidum subsp. pallidum, has now been cultured in vitro continuously for periods exceeding 3 years using a system consisting of coculture with Sf1Ep rabbit epithelial cells in TpCM-2 medium and a low-oxygen environment. In addition, long-term culture of several other syphilis isolates (SS14, Mexico A, UW231B, and UW249B) and the T. pallidum subsp. endemicum Bosnia A strain has been achieved. During in vitro passage, T. pallidum subsp. pallidum exhibited a typical bacterial growth curve with logarithmic and stationary phases. Sf1Ep cells are required for sustained growth and motility; however, high initial Sf1Ep cell numbers resulted in reduced multiplication and survival. Use of Eagle's minimal essential medium as the basal medium was not effective in sustaining growth of T. pallidum subsp. pallidum beyond the first passage, whereas CMRL 1066 or M199 supported long-term culture, confirming that additional nutrients present in these more complex basal media are required for long-term culture. T. pallidum subsp. pallidum growth was dependent upon the presence of fetal bovine serum, with 20% (vol/vol) being the optimal concentration. Omission of reactive oxygen species scavengers dithiothreitol, d-mannitol, or l-histidine did not dramatically affect survival or growth. Additionally, T. pallidum subsp. pallidum can be successfully cultured in a Brewer jar instead of a specialized low-oxygen incubator. Phosphomycin or amphotericin B can be added to the medium to aid in the prevention of bacterial or fungal contamination, respectively. These results help define the parameters of the T. pallidum subsp. pallidum culture system that are required for sustained, long-term survival and multiplication.IMPORTANCE Syphilis is caused by the bacterium Treponema pallidum subsp. pallidum Until recently, this pathogen could only be maintained through infection of rabbits or other animals, making study of this important human pathogen challenging and costly. T. pallidum subsp. pallidum has now been successfully cultured for over 3 years in a tissue culture system using a medium called TpCM-2. Here, we further define the growth requirements of this important human pathogen, promoting a better understanding of the biology of this fastidious organism.
Keywords: Treponema pallidum; bejel; culture; epithelial cells; growth requirements; oxygen; physiology; syphilis.
Copyright © 2021 Edmondson et al.
Figures

Similar articles
-
Long-Term In Vitro Culture of the Syphilis Spirochete Treponema pallidum subsp. pallidum.mBio. 2018 Jun 26;9(3):e01153-18. doi: 10.1128/mBio.01153-18. mBio. 2018. PMID: 29946052 Free PMC article.
-
Procedures for In Vitro Cultivation of Treponema pallidum, the Syphilis Spirochete.J Vis Exp. 2025 Jan 24;(215). doi: 10.3791/66880. J Vis Exp. 2025. PMID: 39927652
-
Cultivation of cottontail rabbit epidermal (Sf1Ep) cells on microcarrier beads and their use for suspension cultivation of Treponema pallidum subsp. pallidum.Appl Environ Microbiol. 1988 Nov;54(11):2862-5. doi: 10.1128/aem.54.11.2862-2865.1988. Appl Environ Microbiol. 1988. PMID: 3063209 Free PMC article.
-
Establishment of the Nichols strain as the type strain of Treponema pallidum.Int J Syst Evol Microbiol. 2025 Feb;75(2):006697. doi: 10.1099/ijsem.0.006697. Int J Syst Evol Microbiol. 2025. PMID: 40014041 Free PMC article. Review.
-
Genetic diversity in Treponema pallidum: implications for pathogenesis, evolution and molecular diagnostics of syphilis and yaws.Infect Genet Evol. 2012 Mar;12(2):191-202. doi: 10.1016/j.meegid.2011.12.001. Epub 2011 Dec 15. Infect Genet Evol. 2012. PMID: 22198325 Free PMC article. Review.
Cited by
-
In Vitro Transformation and Selection of Treponema pallidum subsp. pallidum.Curr Protoc. 2022 Aug;2(8):e507. doi: 10.1002/cpz1.507. Curr Protoc. 2022. PMID: 35976045 Free PMC article.
-
Tp40: a new potential prognostic and diagnostic marker for syphilis.Microbiol Spectr. 2025 Mar 4;13(3):e0279924. doi: 10.1128/spectrum.02799-24. Epub 2025 Feb 11. Microbiol Spectr. 2025. PMID: 39932281 Free PMC article.
-
Design of Epitopes from Treponema pallidum Lipoprotein Antigens for Syphilis Diagnosis and Treatment Prognosis.ACS Infect Dis. 2025 Jun 13;11(6):1606-1622. doi: 10.1021/acsinfecdis.5c00155. Epub 2025 May 23. ACS Infect Dis. 2025. PMID: 40407751 Free PMC article.
-
A novel pan-proteome array for high-throughput profiling of the humoral response to Treponema pallidum.iScience. 2024 Jul 31;27(9):110618. doi: 10.1016/j.isci.2024.110618. eCollection 2024 Sep 20. iScience. 2024. PMID: 39262771 Free PMC article.
-
In Vitro Cultivation of the Syphilis Spirochete Treponema pallidum.Curr Protoc. 2021 Feb;1(2):e44. doi: 10.1002/cpz1.44. Curr Protoc. 2021. PMID: 33599121 Free PMC article.
References
-
- Schaudinn F, Hoffmann E. 1905. Über spirochaetenbefunde im lymphdrüsensaft syphilitischer. Dtsch Med Wochenschr 31:711–714. doi: 10.1055/s-0029-1188114. - DOI
-
- Schaudinn F, Hoffman E. 1905. Vorläufiger bericht über das vorkommen für spirochaeten in syphilitischen krankheitsprodukten und bei papillomen. Arb Gesundh Amt Berlin 22:528–534.
-
- Radolf JD, Lukehart SA (ed). 2006. Pathogenic Treponema: molecular and cellular biology. Caister Academic Press, Hethersett, Norwich, UK.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
