

A Bacterial Toxin Perturbs Intracellular Amino Acid Balance To Induce Persistence
- PMID: 33622732
- PMCID: PMC8545095
- DOI: 10.1128/mBio.03020-20
A Bacterial Toxin Perturbs Intracellular Amino Acid Balance To Induce Persistence
Abstract
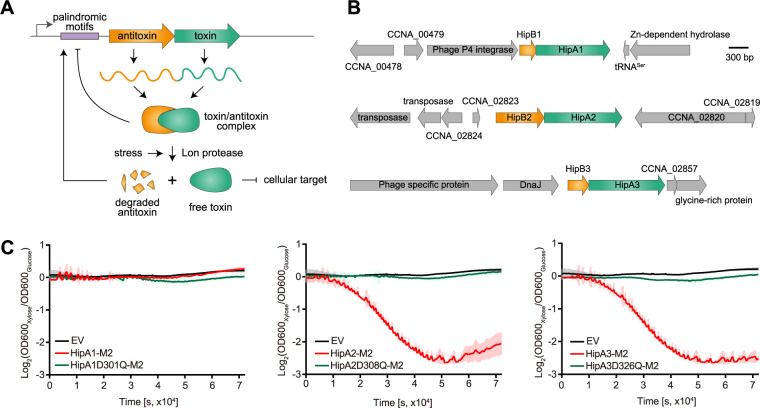
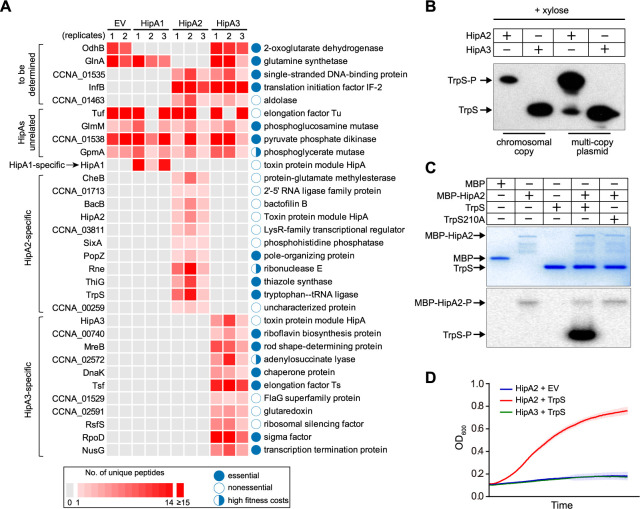
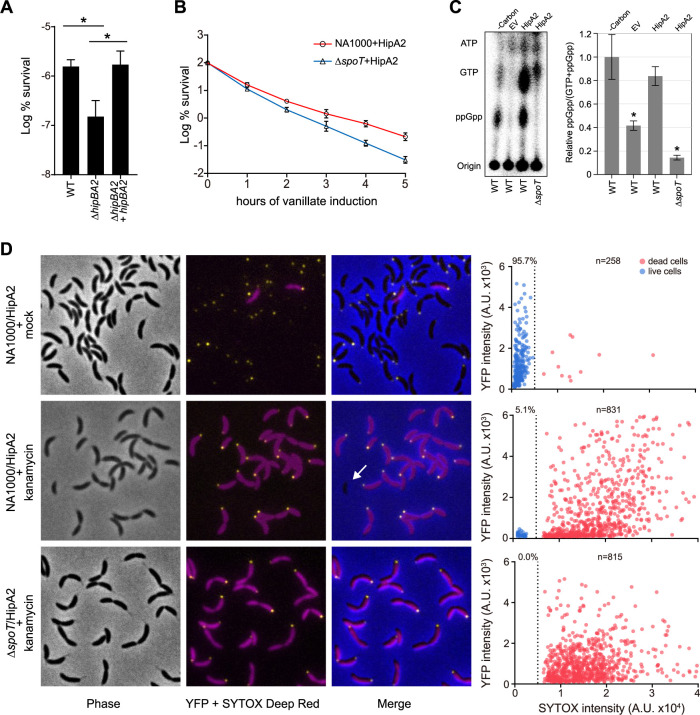
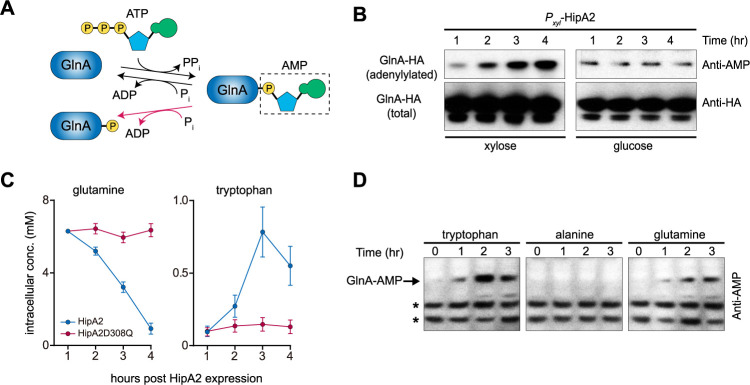
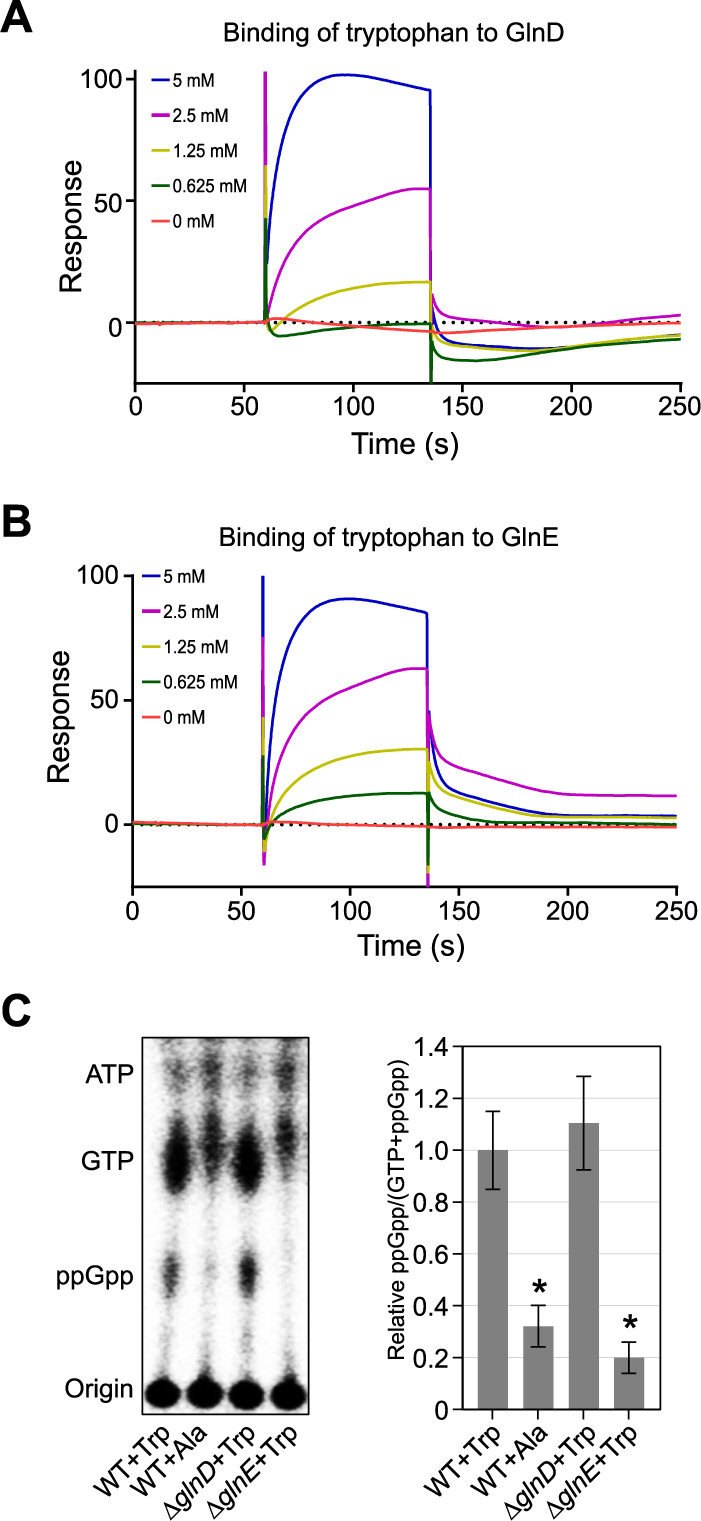
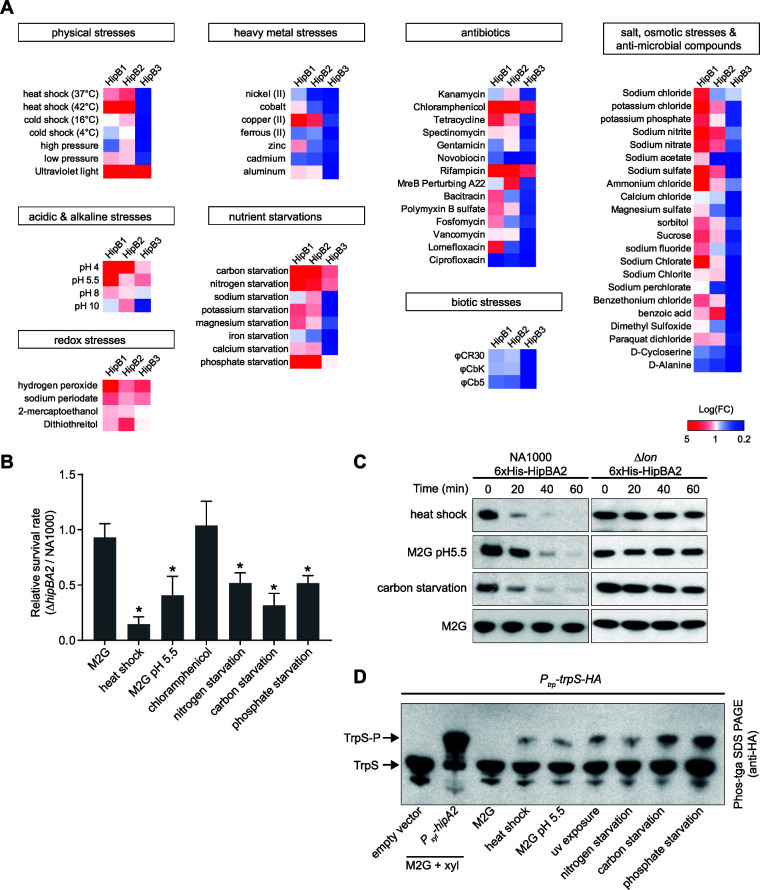
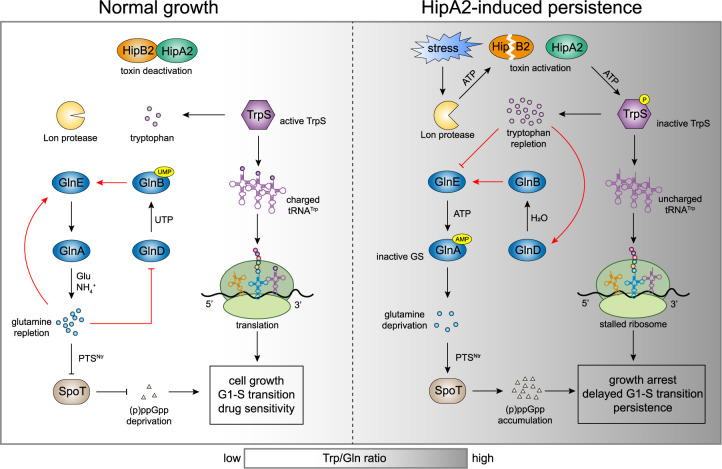
Bacterial cells utilize toxin-antitoxin systems to inhibit self-reproduction, while maintaining viability, when faced with environmental challenges. The activation of the toxin is often coupled to the induction of cellular response pathways, such as the stringent response, in response to multiple stress conditions. Under these conditions, the cell enters a quiescent state referred to as dormancy or persistence. How toxin activation triggers persistence and induces a systemic stress response in the alphaproteobacteria remains unclear. Here, we report that in Caulobacter, a hipA2-encoded bacterial toxin contributes to bacterial persistence by manipulating intracellular amino acid balance. HipA2 is a serine/threonine kinase that deactivates tryptophanyl-tRNA synthetase by phosphorylation, leading to stalled protein synthesis and the accumulation of free tryptophan. An increased level of tryptophan allosterically activates the adenylyltransferase activity of GlnE that, in turn, deactivates glutamine synthetase GlnA by adenylylation. The inactivation of GlnA promotes the deprivation of glutamine in the cell, which triggers a stringent response. By screening 69 stress conditions, we find that HipBA2 responds to multiple stress signals through the proteolysis of HipB2 antitoxin by the Lon protease and the release of active HipA2 kinase, revealing a molecular mechanism that allows disparate stress conditions to be sensed and funneled into a single response pathway.IMPORTANCE To overcome various environmental challenges, bacterial cells can enter a physiologically quiescent state, known as dormancy or persistence, which balances growth and viability. In this study, we report a new mechanism by which a toxin-antitoxin system responds to harsh environmental conditions or nutrient deprivation by orchestrating a dormant state while preserving viability. The hipA2-encoded kinase functions as a toxin in Caulobacter, inducing bacterial persistence by disturbing the intracellular tryptophan-glutamine balance. A nitrogen regulatory circuit can be regulated by the intracellular level of tryptophan, which mimics the allosteric role of glutamine in this feedback loop. The HipBA2 module senses different types of stress conditions by increasing the intracellular level of tryptophan, which in turn breaks the tryptophan-glutamine balance and induces glutamine deprivation. Our results reveal a molecular mechanism that allows disparate environmental challenges to converge on a common pathway that results in a dormant state.
Keywords: Caulobacter crescentus; amino acid balance; bacterial toxin; persistence; stringent response.
Copyright © 2021 Zhou et al.
Figures
Similar articles
-
Serine-Threonine Kinases Encoded by Split hipA Homologs Inhibit Tryptophanyl-tRNA Synthetase.mBio. 2019 Jun 18;10(3):e01138-19. doi: 10.1128/mBio.01138-19. mBio. 2019. PMID: 31213559 Free PMC article.
-
hipBA toxin-antitoxin systems mediate persistence in Caulobacter crescentus.Sci Rep. 2020 Feb 18;10(1):2865. doi: 10.1038/s41598-020-59283-x. Sci Rep. 2020. PMID: 32071324 Free PMC article.
-
Homologous VapC Toxins Inhibit Translation and Cell Growth by Sequence-Specific Cleavage of tRNAfMet.J Bacteriol. 2018 Jan 10;200(3):e00582-17. doi: 10.1128/JB.00582-17. Print 2018 Feb 1. J Bacteriol. 2018. PMID: 29109187 Free PMC article.
-
RNA-based regulation in type I toxin-antitoxin systems and its implication for bacterial persistence.Curr Genet. 2017 Dec;63(6):1011-1016. doi: 10.1007/s00294-017-0710-y. Epub 2017 May 30. Curr Genet. 2017. PMID: 28560584 Free PMC article. Review.
-
Toxins targeting transfer RNAs: Translation inhibition by bacterial toxin-antitoxin systems.Wiley Interdiscip Rev RNA. 2019 Jan;10(1):e1506. doi: 10.1002/wrna.1506. Epub 2018 Sep 16. Wiley Interdiscip Rev RNA. 2019. PMID: 30296016 Free PMC article. Review.
Cited by
-
The Bro-Xre toxin-antitoxin modules in Weissella cibaria: inducing persister cells to escape tetracycline stress by disrupting metabolism.Front Microbiol. 2024 Nov 29;15:1505841. doi: 10.3389/fmicb.2024.1505841. eCollection 2024. Front Microbiol. 2024. PMID: 39678910 Free PMC article.
-
Bacterial cell differentiation enables population level survival strategies.mBio. 2024 Jun 12;15(6):e0075824. doi: 10.1128/mbio.00758-24. Epub 2024 May 21. mBio. 2024. PMID: 38771034 Free PMC article. Review.
-
Deacylated tRNA Accumulation Is a Trigger for Bacterial Antibiotic Persistence Independent of the Stringent Response.mBio. 2021 Jun 29;12(3):e0113221. doi: 10.1128/mBio.01132-21. Epub 2021 Jun 15. mBio. 2021. PMID: 34126764 Free PMC article.
-
The plasmid-borne hipBA operon of Klebsiella michiganensis encodes a potent plasmid stabilization system.J Appl Microbiol. 2024 Oct 3;135(10):lxae246. doi: 10.1093/jambio/lxae246. J Appl Microbiol. 2024. PMID: 39304528 Free PMC article.
-
Contribution of Toxin-Antitoxin Systems to Adherent-Invasive E. coli Pathogenesis.Microorganisms. 2024 Jun 6;12(6):1158. doi: 10.3390/microorganisms12061158. Microorganisms. 2024. PMID: 38930540 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
