

Functional connections between cell cycle and proteostasis in the regulation of Candida albicans morphogenesis
- PMID: 33626353
- PMCID: PMC7971348
- DOI: 10.1016/j.celrep.2021.108781
Functional connections between cell cycle and proteostasis in the regulation of Candida albicans morphogenesis
Abstract
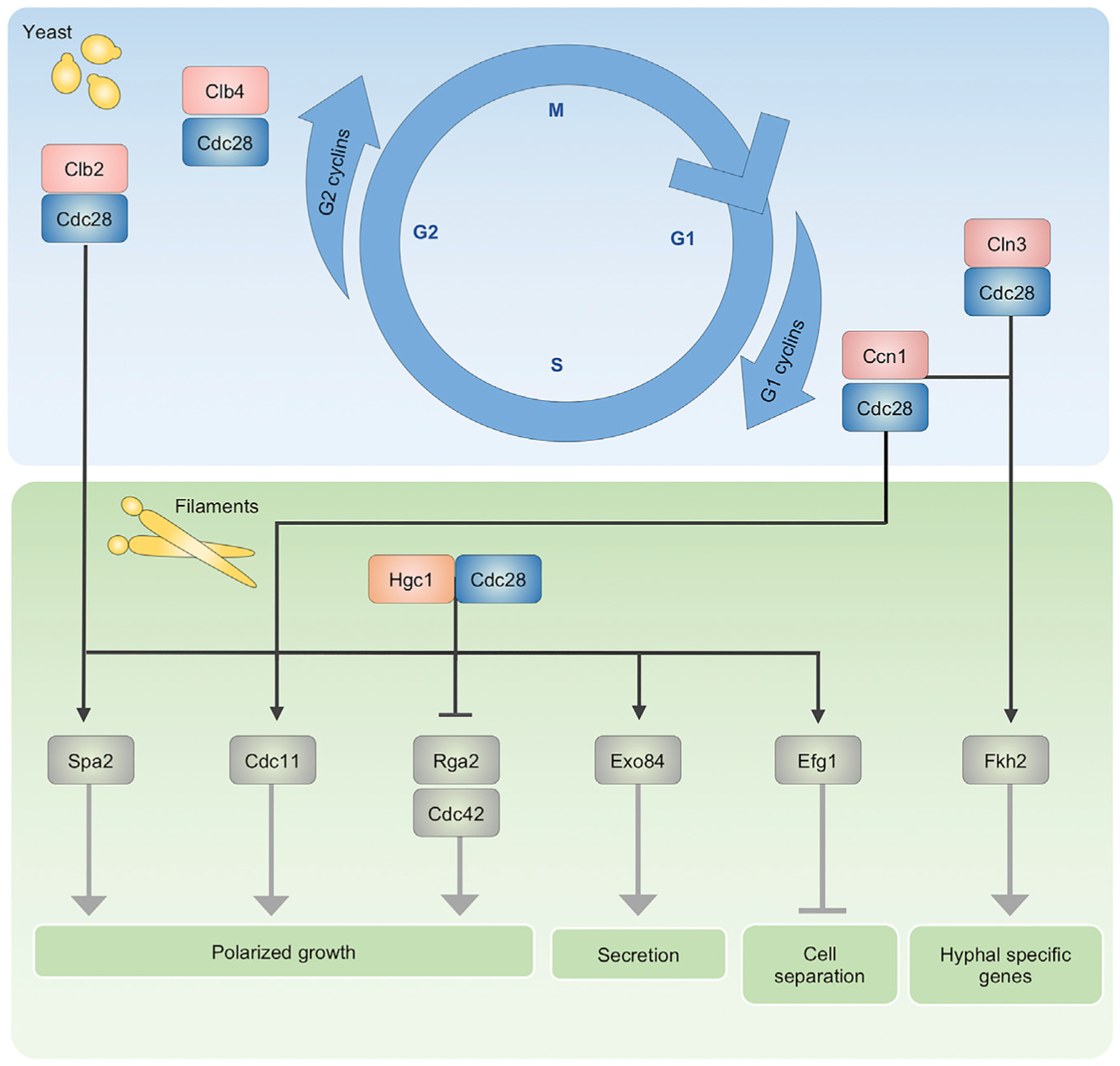
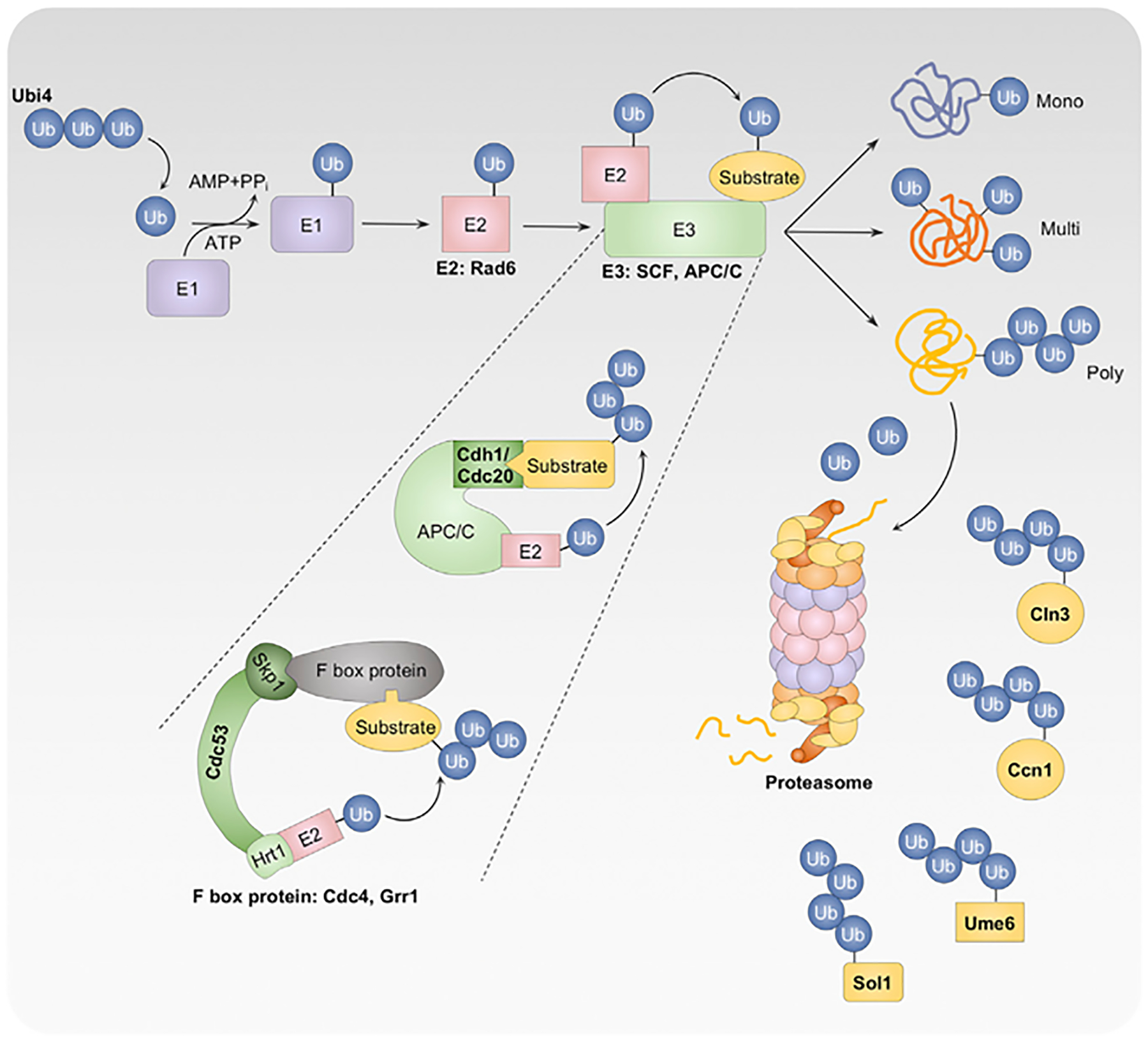
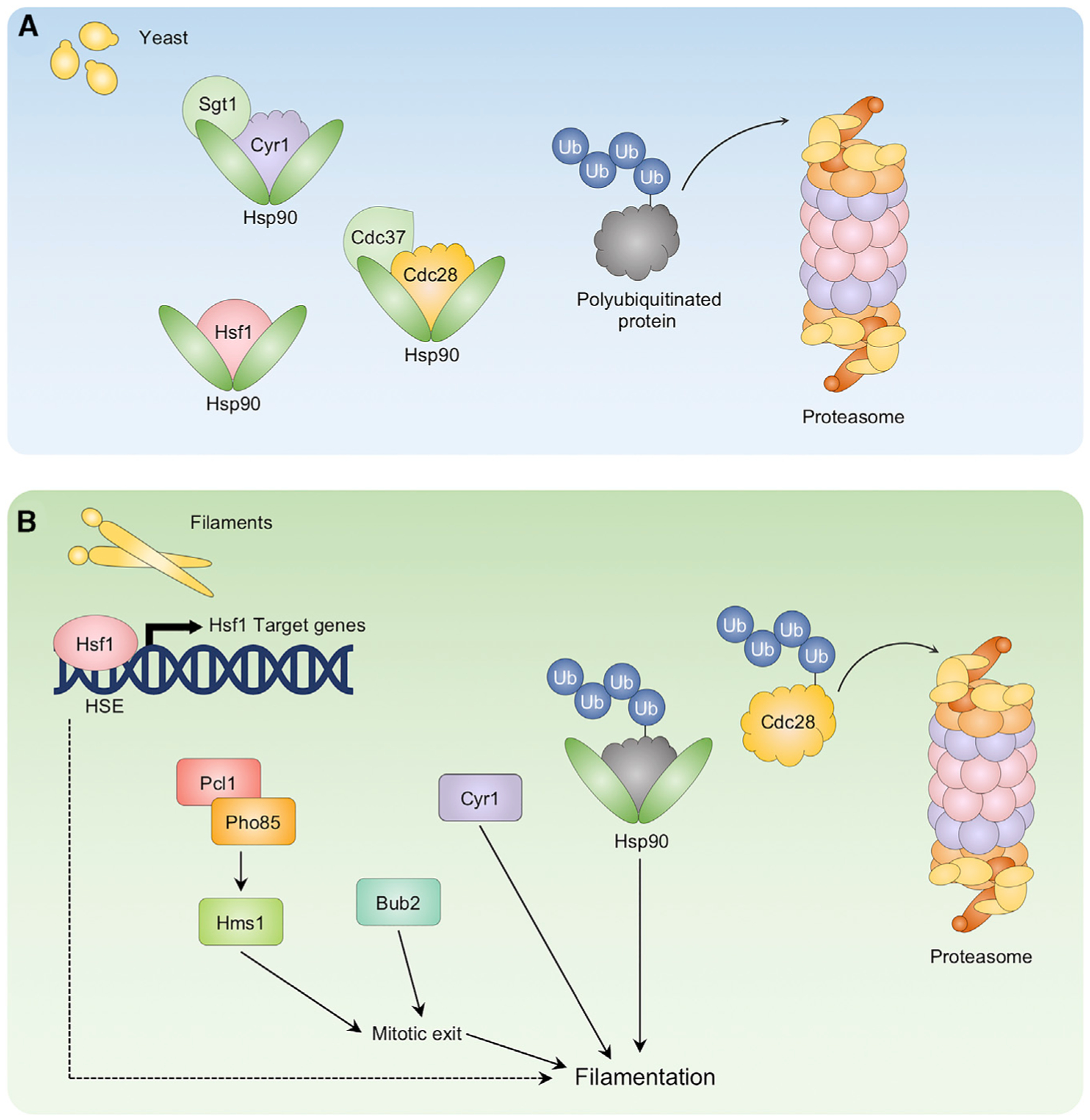
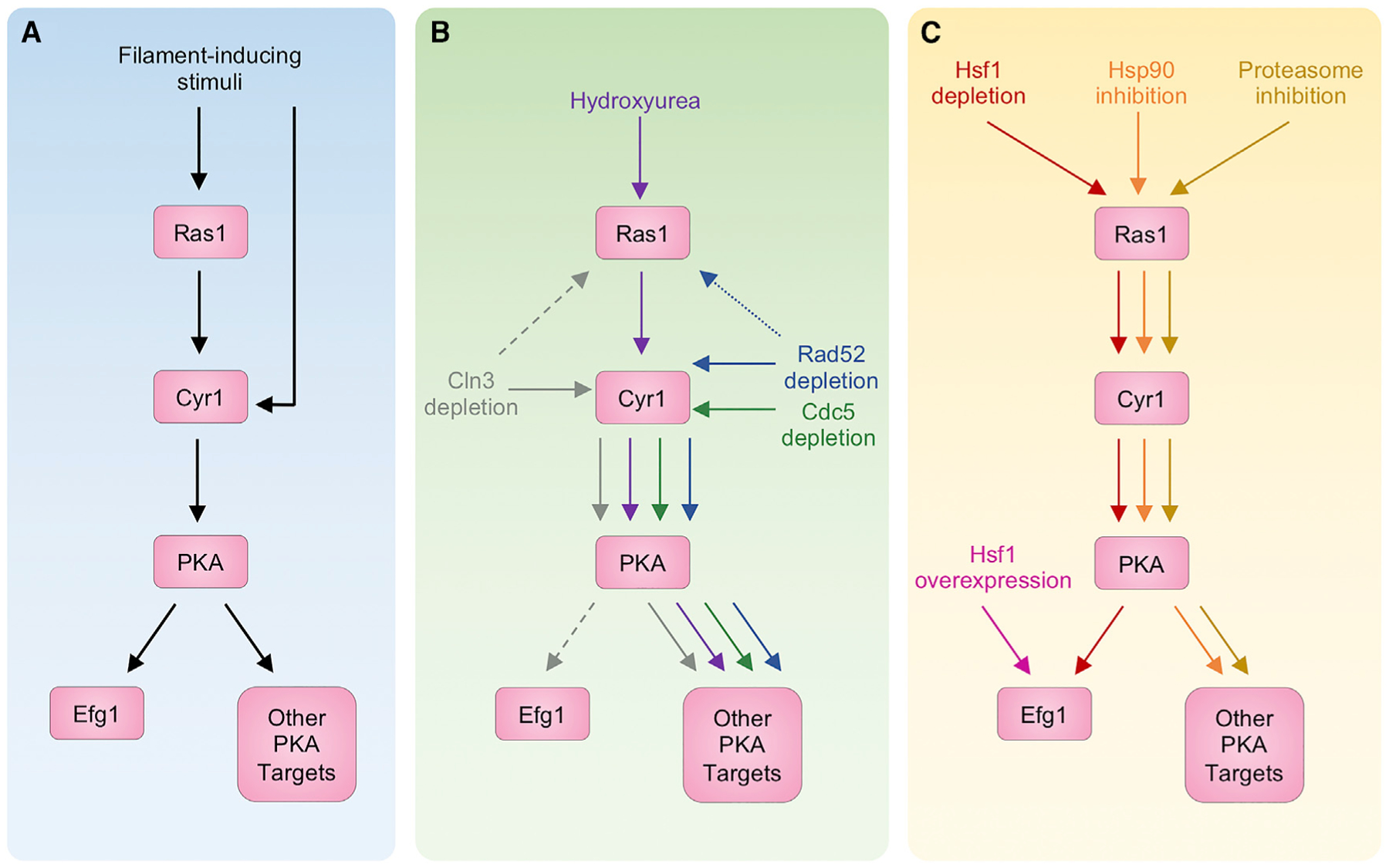
Morphological plasticity is a key virulence trait for many fungal pathogens. For the opportunistic fungal pathogen Candida albicans, transitions among yeast, pseudohyphal, and hyphal forms are critical for virulence, because the morphotypes play distinct roles in the infection process. C. albicans morphogenesis is induced in response to many host-relevant conditions and is regulated by complex signaling pathways and cellular processes. Perturbation of either cell-cycle progression or protein homeostasis induces C. albicans filamentation, demonstrating that these processes play a key role in morphogenetic control. Regulators such as cyclin-dependent kinases, checkpoint proteins, the proteasome, the heat shock protein Hsp90, and the heat shock transcription factor Hsf1 all influence morphogenesis, often through interconnected effects on the cell cycle and proteostasis. This review highlights the major cell-cycle and proteostasis regulators that modulate morphogenesis and discusses how these two processes intersect to regulate this key virulence trait.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.E.C. is a cofounder and shareholder in Bright Angel Therapeutics, a platform company for development of novel antifungal therapeutics. L.E.C. is also a consultant for Boragen, a small-molecule development company focused on leveraging the unique chemical properties of boron chemistry for crop protection and animal health.
Figures
References
-
- Adams J (2004). The proteasome in cell-cycle regulation. In Proteasome Inhibitors in Cancer Therapy, Adams J, ed. (Humana Press; ), pp. 77–84.
-
- Andaluz E, Ciudad T, Gómez-Raja J, Calderone R, and Larriba G (2006). Rad52 depletion in Candida albicans triggers both the DNA-damage checkpoint and filamentation accompanied by but independent of expression of hypha-specific genes. Mol. Microbiol 59, 1452–1472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
