

Functional packaging of seeds
- PMID: 33629369
- PMCID: PMC8252473
- DOI: 10.1111/nph.17299
Functional packaging of seeds
Abstract
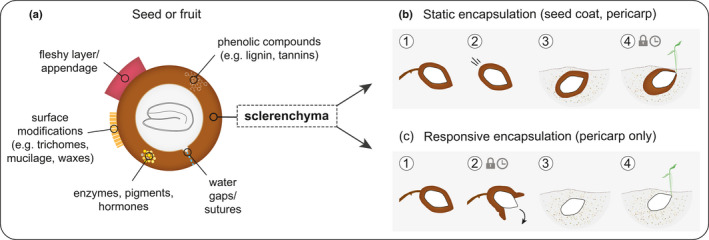
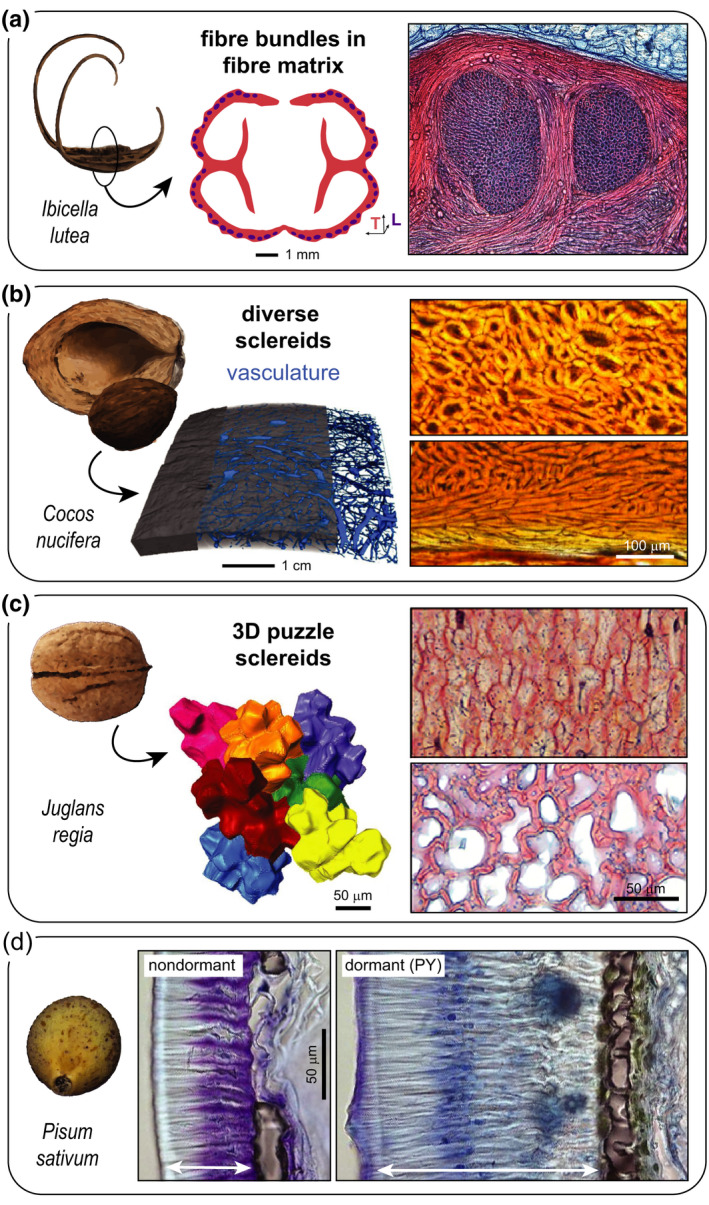
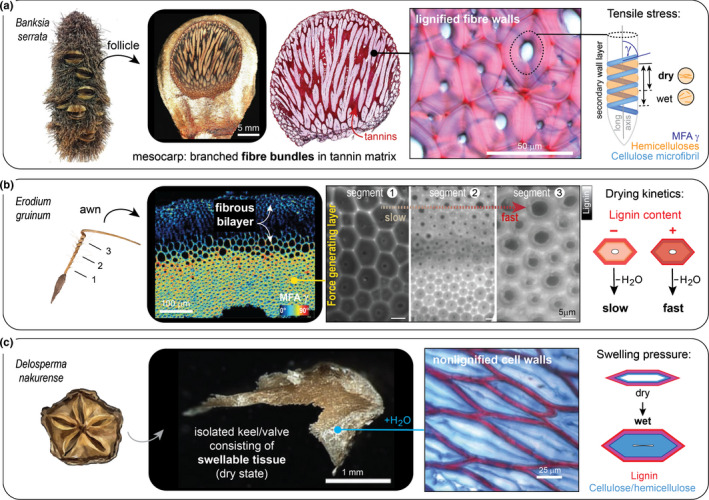
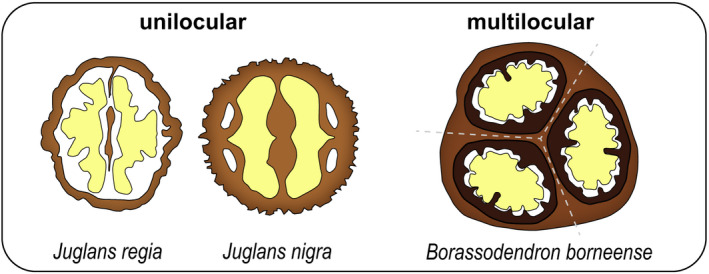
The encapsulation of seeds in hard coats and fruit walls (pericarp layers) fulfils protective and dispersal functions in many plant families. In angiosperms, packaging structures possess a remarkable range of different morphologies and functionalities, as illustrated by thermo and hygro-responsive seed pods and appendages, as well as mechanically strong and water-impermeable shells. Key to these different functionalities are characteristic structural arrangements and chemical modifications of the underlying sclerenchymatous tissues. Although many ecological aspects of hard seed encapsulation have been well documented, a detailed understanding of the relationship between tissue structure and function only recently started to emerge, especially in the context of environmentally driven fruit opening and seed dispersal (responsive encapsulations) and the outstanding durability of some seed coats and indehiscent fruits (static encapsulations). In this review, we focus on the tissue properties of these two systems, with particular consideration of water interactions, mechanical resistance, and force generation. Common principles, as well as unique adaptations, are discussed in different plant species. Understanding how plants integrate a broad range of functions and properties for seed protection during storage and dispersal plays a central role for seed conservation, population dynamics, and plant-based material developments.
Keywords: diaspore adaptations; hard plant shells; physical dormancy; sclerenchyma; seed storage and protection; serotiny; tissue mechanics.
© 2021 The Authors New Phytologist © 2021 New Phytologist Foundation.
Figures
Similar articles
-
Dispersal biophysics and adaptive significance of dimorphic diaspores in the annual Aethionema arabicum (Brassicaceae).New Phytol. 2019 Feb;221(3):1434-1446. doi: 10.1111/nph.15490. Epub 2018 Oct 25. New Phytol. 2019. PMID: 30230555 Free PMC article.
-
A water drop-shaped slingshot in plants: geometry and mechanics in the explosive seed dispersal of Orixa japonica (Rutaceae).Ann Bot. 2021 May 7;127(6):765-774. doi: 10.1093/aob/mcab017. Ann Bot. 2021. PMID: 33608717 Free PMC article.
-
The effect of gut passage by waterbirds on the seed coat and pericarp of diaspores lacking "external flesh": Evidence for widespread adaptation to endozoochory in angiosperms.PLoS One. 2019 Dec 19;14(12):e0226551. doi: 10.1371/journal.pone.0226551. eCollection 2019. PLoS One. 2019. PMID: 31856213 Free PMC article.
-
Seeds-An evolutionary innovation underlying reproductive success in flowering plants.Curr Top Dev Biol. 2019;131:605-642. doi: 10.1016/bs.ctdb.2018.11.017. Epub 2018 Dec 23. Curr Top Dev Biol. 2019. PMID: 30612632 Review.
-
Dead but Not Dead End: Multifunctional Role of Dead Organs Enclosing Embryos in Seed Biology.Int J Mol Sci. 2020 Oct 28;21(21):8024. doi: 10.3390/ijms21218024. Int J Mol Sci. 2020. PMID: 33126660 Free PMC article. Review.
Cited by
-
Seed-to-Seedling Transition: Novel Aspects.Plants (Basel). 2022 Jul 30;11(15):1988. doi: 10.3390/plants11151988. Plants (Basel). 2022. PMID: 35956466 Free PMC article.
-
Ginkgo seed shell provides a unique model for bioinspired design.Proc Natl Acad Sci U S A. 2022 Dec 6;119(49):e2211458119. doi: 10.1073/pnas.2211458119. Epub 2022 Nov 28. Proc Natl Acad Sci U S A. 2022. PMID: 36442101 Free PMC article.
-
Biomimicking Nature-Inspired Design Structures-An Experimental and Simulation Approach Using Additive Manufacturing.Biomimetics (Basel). 2022 Nov 3;7(4):186. doi: 10.3390/biomimetics7040186. Biomimetics (Basel). 2022. PMID: 36412714 Free PMC article.
-
Fruit dehiscence mechanism and release of dimorphic seeds with different germination properties in Commelina erecta.Protoplasma. 2024 Mar;261(2):377-393. doi: 10.1007/s00709-023-01904-z. Epub 2023 Nov 1. Protoplasma. 2024. PMID: 37910229
-
Fire-released seed dormancy - a global synthesis.Biol Rev Camb Philos Soc. 2022 Aug;97(4):1612-1639. doi: 10.1111/brv.12855. Epub 2022 Apr 6. Biol Rev Camb Philos Soc. 2022. PMID: 35384243 Free PMC article.
References
-
- Abraham Y, Dong Y, Aharoni A, Elbaum R. 2018. Mapping of cell wall aromatic moieties and their effect on hygroscopic movement in the awns of stork’s bill. Cellulose 25: 3827–3841.
-
- Abraham Y, Elbaum R. 2013. Hygroscopic movements in Geraniaceae: the structural variations that are responsible for coiling or bending. New Phytologist 199: 584–594. - PubMed
-
- Armon S, Efrati E, Kupferman R, Sharon E. 2011. Geometry and mechanics in the opening of chiral seed pods. Science 333: 1726–1730. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
