

An aspartyl protease-mediated cleavage regulates structure and function of a flavodoxin-like protein and aids oxidative stress survival
- PMID: 33630938
- PMCID: PMC7943015
- DOI: 10.1371/journal.ppat.1009355
An aspartyl protease-mediated cleavage regulates structure and function of a flavodoxin-like protein and aids oxidative stress survival
Abstract
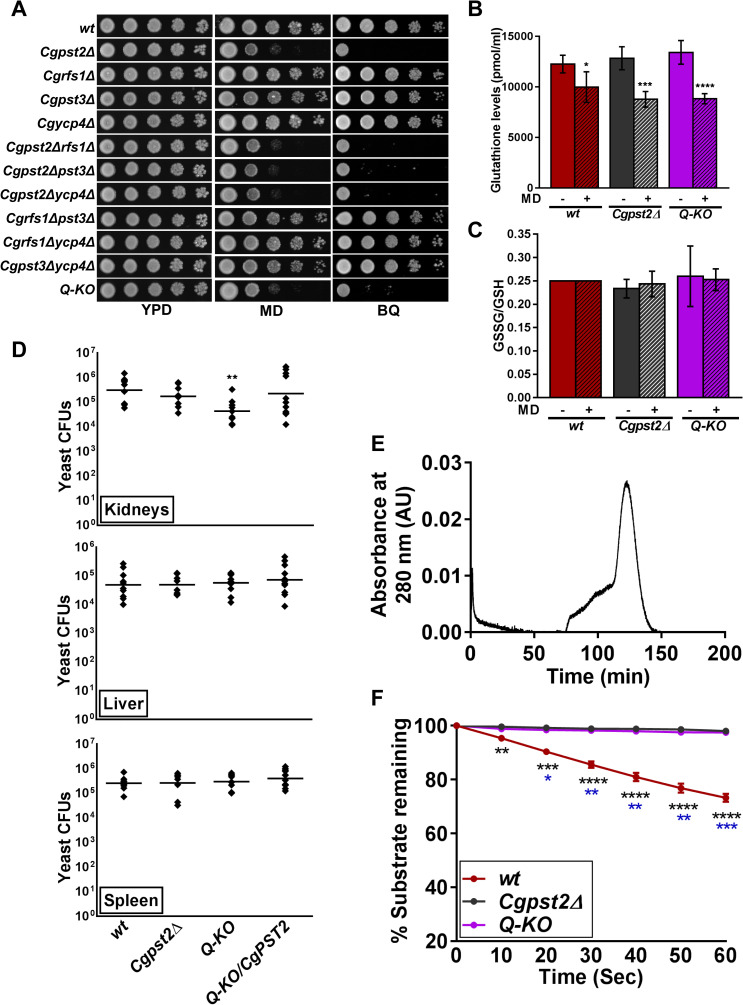
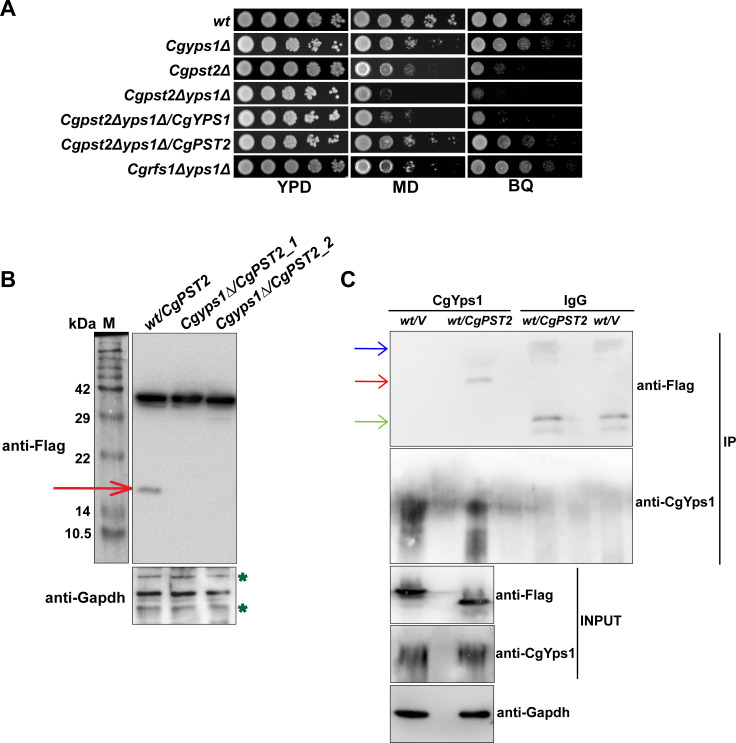
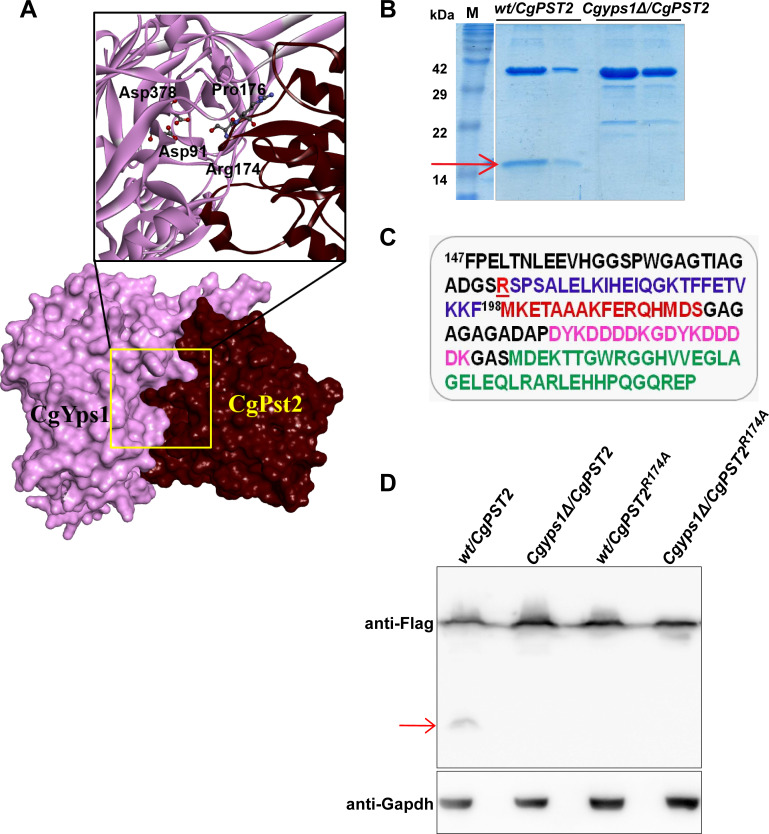
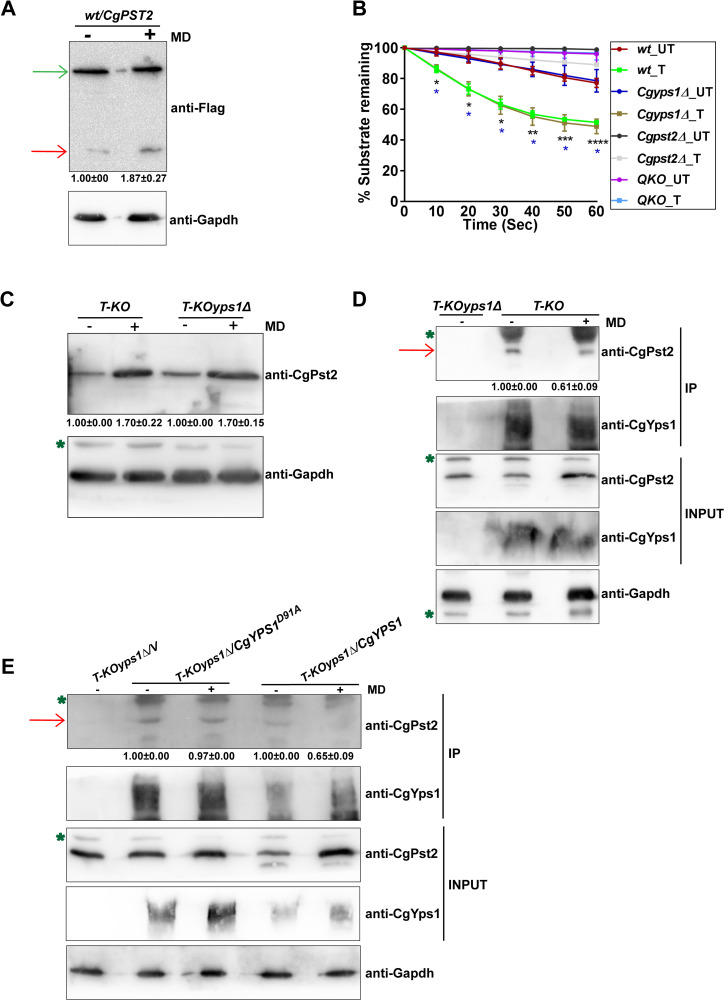
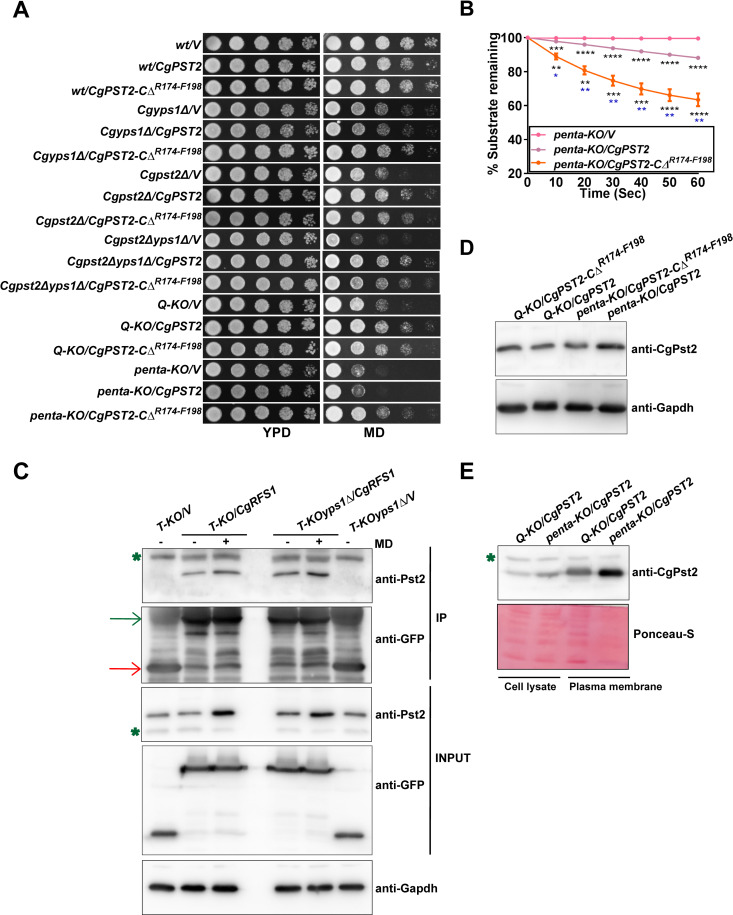
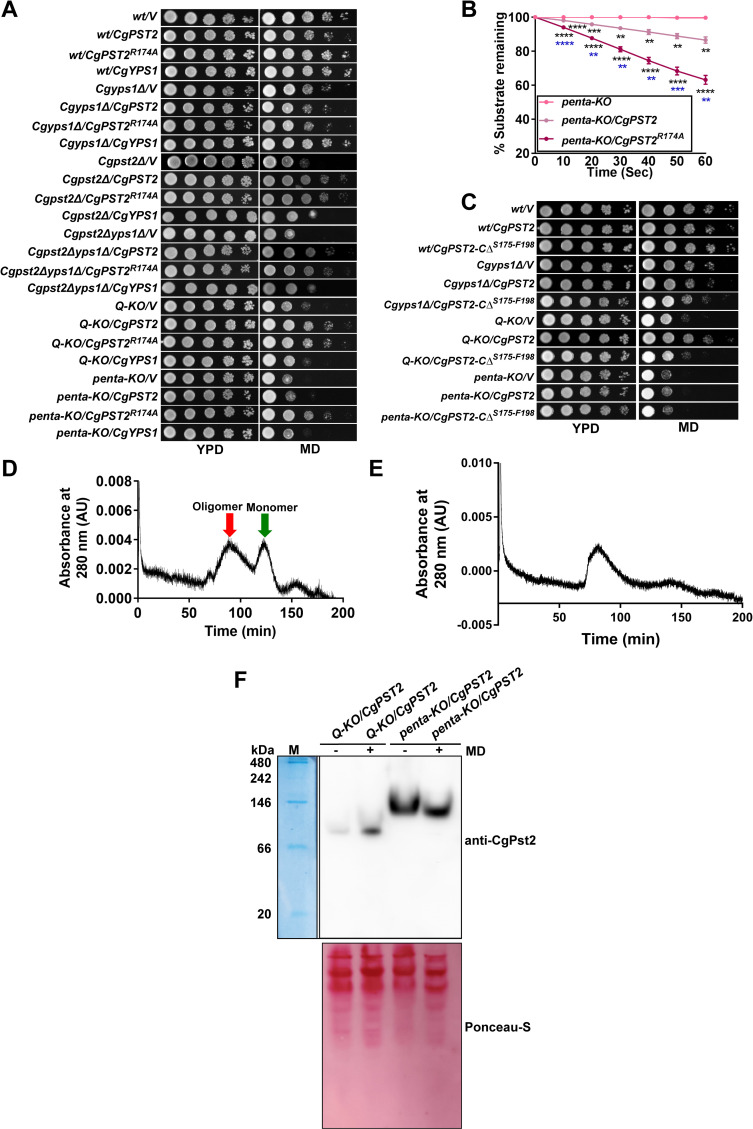
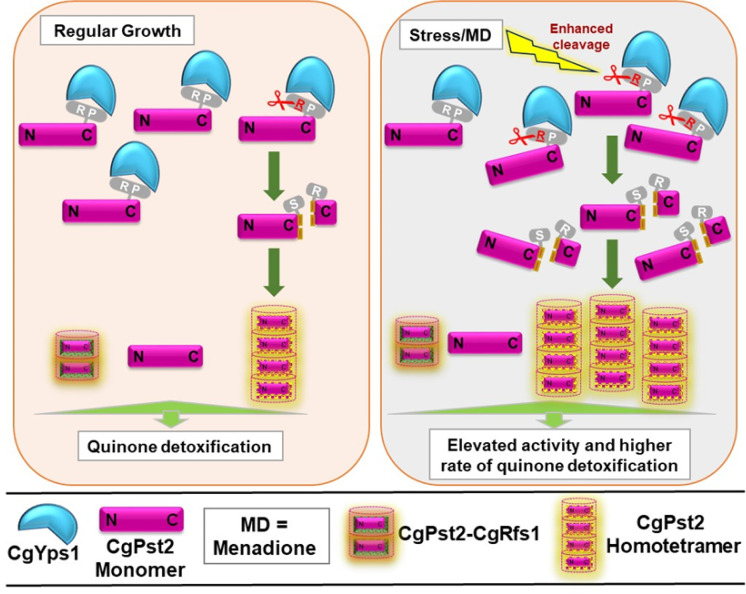
A family of eleven glycosylphosphatidylinositol-anchored aspartyl proteases, commonly referred to as CgYapsins, regulate a myriad of cellular processes in the pathogenic yeast Candida glabrata, but their protein targets are largely unknown. Here, using the immunoprecipitation-mass spectrometry approach, we identify the flavodoxin-like protein (Fld-LP), CgPst2, to be an interactor of one of the aspartyl protease CgYps1. We also report the presence of four Fld-LPs in C. glabrata, which are required for survival in kidneys in the murine model of systemic candidiasis. We further demonstrated that of four Fld-LPs, CgPst2 was solely required for menadione detoxification. CgPst2 was found to form homo-oligomers, and contribute to cellular NADH:quinone oxidoreductase activity. CgYps1 cleaved CgPst2 at the C-terminus, and this cleavage was pivotal to oligomerization, activity and function of CgPst2. The arginine-174 residue in CgPst2 was essential for CgYps1-mediated cleavage, with alanine substitution of the arginine-174 residue also leading to elevated activity and oligomerization of CgPst2. Finally, we demonstrate that menadione treatment led to increased CgPst2 and CgYps1 protein levels, diminished CgYps1-CgPst2 interaction, and enhanced CgPst2 cleavage and activity, thereby implicating CgYps1 in activating CgPst2. Altogether, our findings of proteolytic cleavage as a key regulatory determinant of CgPst2, which belongs to the family of highly conserved, electron-carrier flavodoxin-fold-containing proteins, constituting cellular oxidative stress defense system in diverse organisms, unveil a hidden regulatory layer of environmental stress response mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
