

Protein Complexes Form a Basis for Complex Hybrid Incompatibility
- PMID: 33633780
- PMCID: PMC7900514
- DOI: 10.3389/fgene.2021.609766
Protein Complexes Form a Basis for Complex Hybrid Incompatibility
Abstract
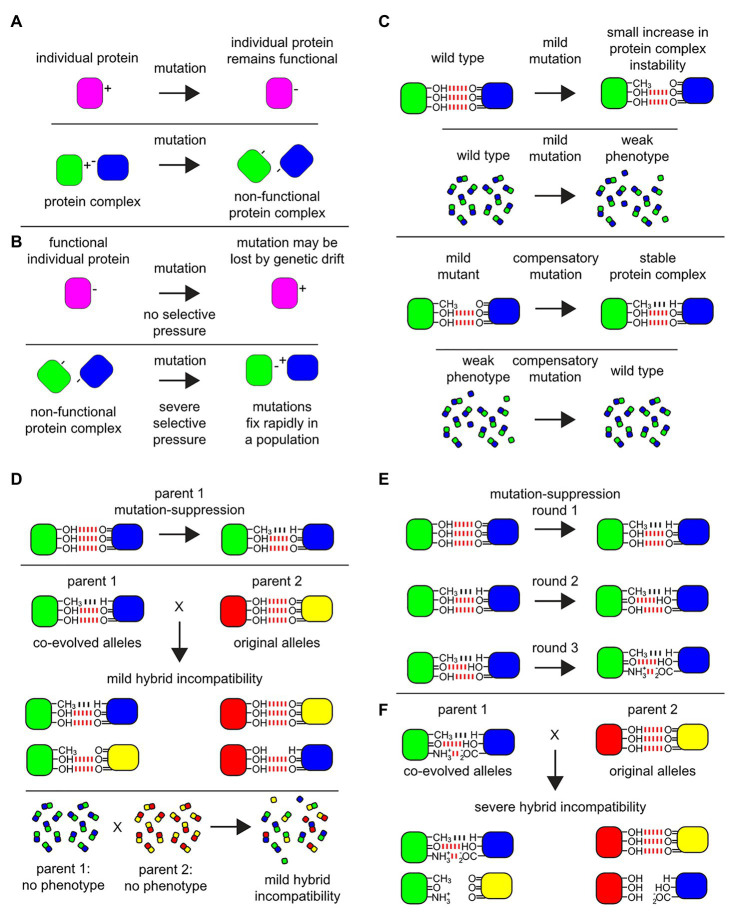
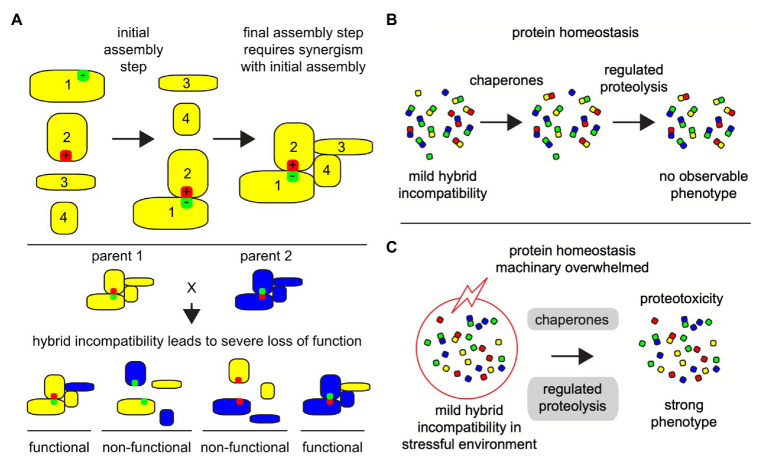
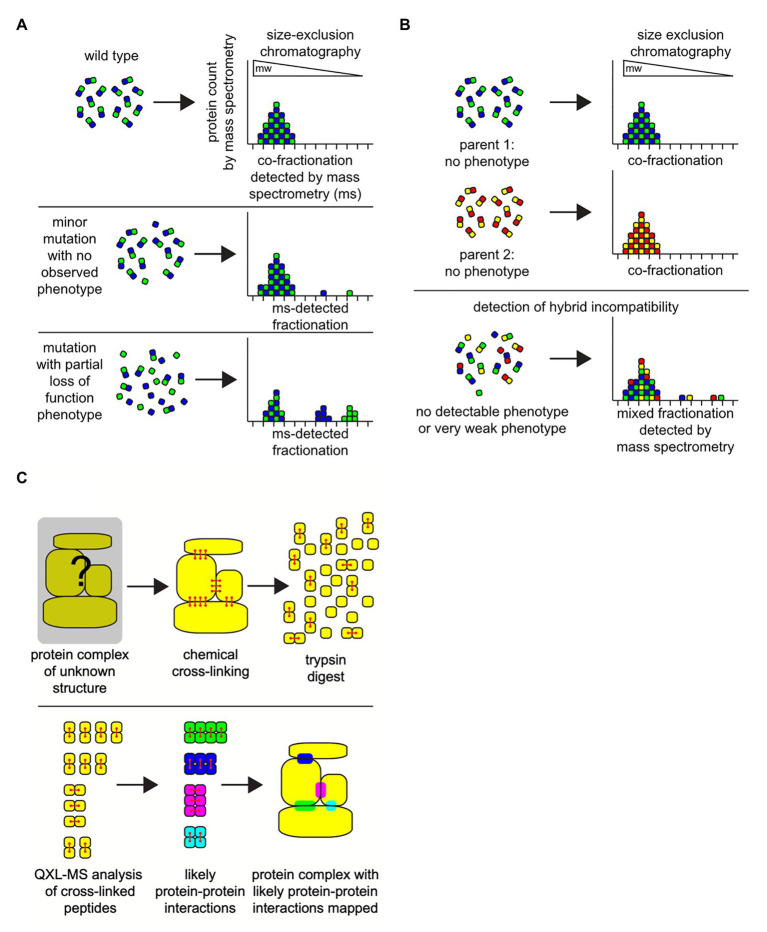
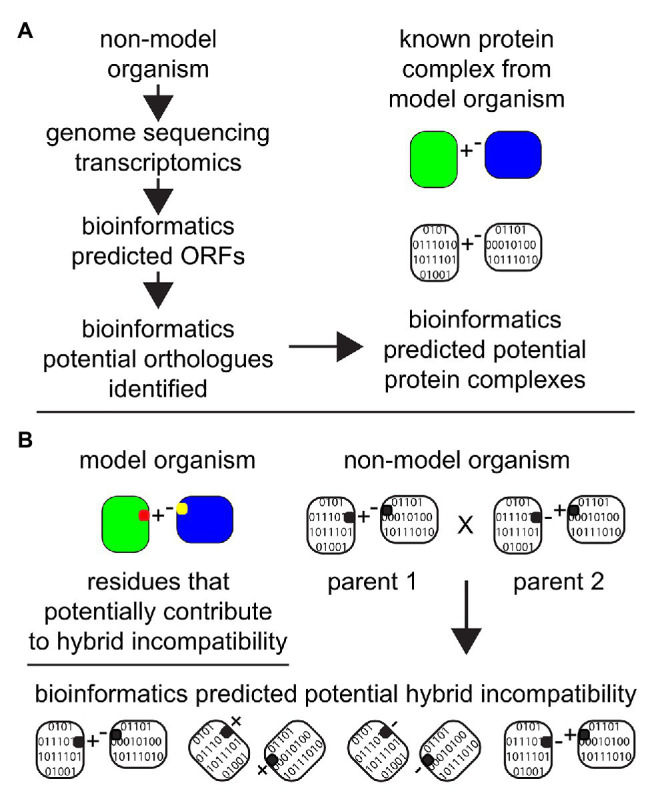
Proteins are the workhorses of the cell and execute many of their functions by interacting with other proteins forming protein complexes. Multi-protein complexes are an admixture of subunits, change their interaction partners, and modulate their functions and cellular physiology in response to environmental changes. When two species mate, the hybrid offspring are usually inviable or sterile because of large-scale differences in the genetic makeup between the two parents causing incompatible genetic interactions. Such reciprocal-sign epistasis between inter-specific alleles is not limited to incompatible interactions between just one gene pair; and, usually involves multiple genes. Many of these multi-locus incompatibilities show visible defects, only in the presence of all the interactions, making it hard to characterize. Understanding the dynamics of protein-protein interactions (PPIs) leading to multi-protein complexes is better suited to characterize multi-locus incompatibilities, compared to studying them with traditional approaches of genetics and molecular biology. The advances in omics technologies, which includes genomics, transcriptomics, and proteomics can help achieve this end. This is especially relevant when studying non-model organisms. Here, we discuss the recent progress in the understanding of hybrid genetic incompatibility; omics technologies, and how together they have helped in characterizing protein complexes and in turn multi-locus incompatibilities. We also review advances in bioinformatic techniques suitable for this purpose and propose directions for leveraging the knowledge gained from model-organisms to identify genetic incompatibilities in non-model organisms.
Keywords: bioinformatics; evolution; hybrid incompatibility; proteins; proteomics; speciation.
Copyright © 2021 Swamy, Schuyler and Leu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Detection of genetic incompatibilities in non-model systems using simple genetic markers: hybrid breakdown in the haplodiploid spider mite Tetranychus evansi.Heredity (Edinb). 2017 Apr;118(4):311-321. doi: 10.1038/hdy.2016.103. Epub 2016 Oct 26. Heredity (Edinb). 2017. PMID: 27782117 Free PMC article.
-
Proteotoxicity caused by perturbed protein complexes underlies hybrid incompatibility in yeast.Nat Commun. 2022 Jul 29;13(1):4394. doi: 10.1038/s41467-022-32107-4. Nat Commun. 2022. PMID: 35906261 Free PMC article.
-
Genomic mechanisms and consequences of diverse postzygotic barriers between monkeyflower species.Genetics. 2023 Nov 1;225(3):iyad156. doi: 10.1093/genetics/iyad156. Genetics. 2023. PMID: 37603838
-
Interspecific hybrids of dwarf hamsters and Phasianidae birds as animal models for studying the genetic and developmental basis of hybrid incompatibility.Genes Genet Syst. 2016 Oct 13;91(2):63-75. doi: 10.1266/ggs.16-00022. Epub 2016 Sep 15. Genes Genet Syst. 2016. PMID: 27628130 Review.
-
Making a Murderer: The Evolutionary Framing of Hybrid Gamete-Killers.Trends Genet. 2019 Apr;35(4):245-252. doi: 10.1016/j.tig.2019.01.004. Epub 2019 Feb 28. Trends Genet. 2019. PMID: 30826132 Review.
Cited by
-
Tick-Tock: Cancer Cell Division Cycle Clocks Strike Midnight.Int J Mol Sci. 2025 Jun 29;26(13):6274. doi: 10.3390/ijms26136274. Int J Mol Sci. 2025. PMID: 40650052 Free PMC article. Review.
-
Hybridization led to a rewired pluripotency network in the allotetraploid Xenopus laevis.Elife. 2023 Oct 3;12:e83952. doi: 10.7554/eLife.83952. Elife. 2023. PMID: 37787392 Free PMC article.
-
Adaptive protein coevolution preserves telomere integrity.bioRxiv [Preprint]. 2024 Nov 11:2024.11.11.623029. doi: 10.1101/2024.11.11.623029. bioRxiv. 2024. PMID: 39605578 Free PMC article. Preprint.
-
Overview of methods for characterization and visualization of a protein-protein interaction network in a multi-omics integration context.Front Mol Biosci. 2022 Sep 8;9:962799. doi: 10.3389/fmolb.2022.962799. eCollection 2022. Front Mol Biosci. 2022. PMID: 36158572 Free PMC article. Review.
-
Glypican-3, Vascular Endothelial Growth Factor and Golgi Protein-73 for Differentiation between Liver Cirrhosis and Hepatocellular Carcinoma.Asian Pac J Cancer Prev. 2023 Feb 1;24(2):497-507. doi: 10.31557/APJCP.2023.24.2.497. Asian Pac J Cancer Prev. 2023. PMID: 36853298 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources