

Nitric Oxide in the Control of the in vitro Proliferation and Differentiation of Human Hematopoietic Stem and Progenitor Cells
- PMID: 33634102
- PMCID: PMC7900502
- DOI: 10.3389/fcell.2020.610369
Nitric Oxide in the Control of the in vitro Proliferation and Differentiation of Human Hematopoietic Stem and Progenitor Cells
Abstract
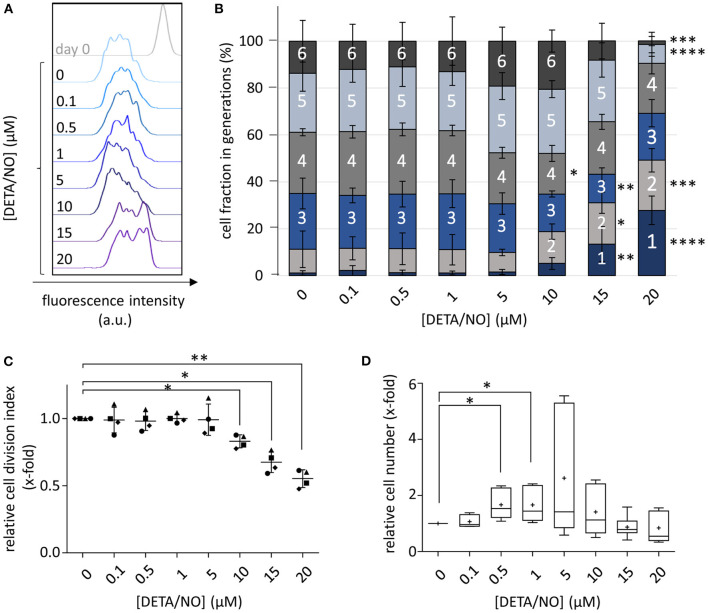
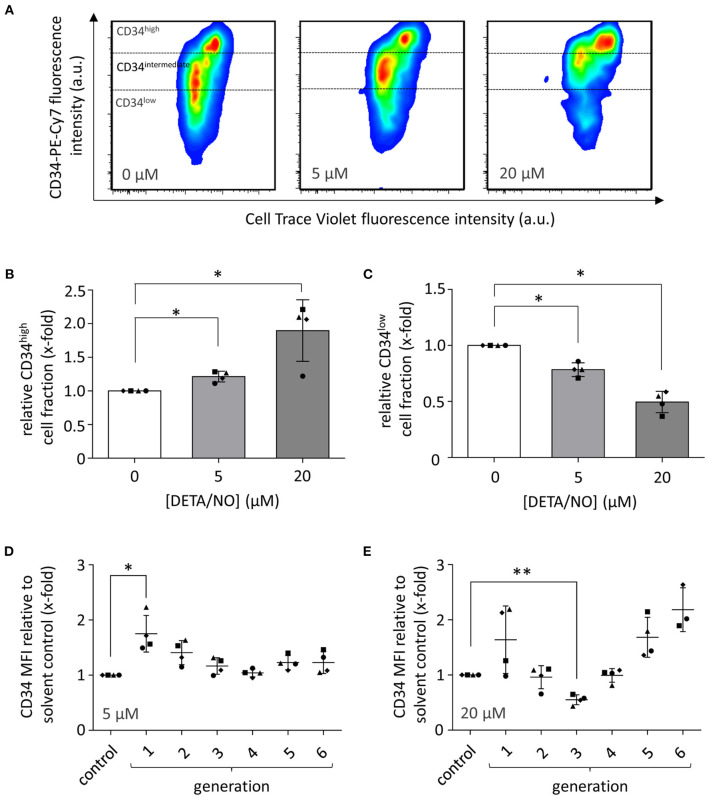
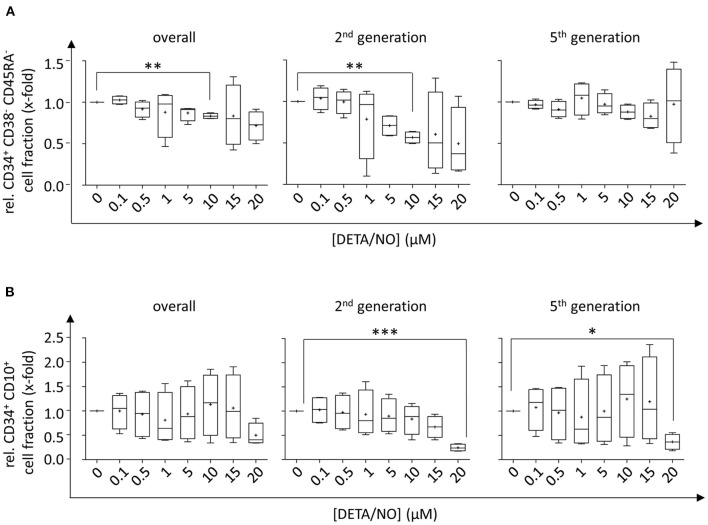
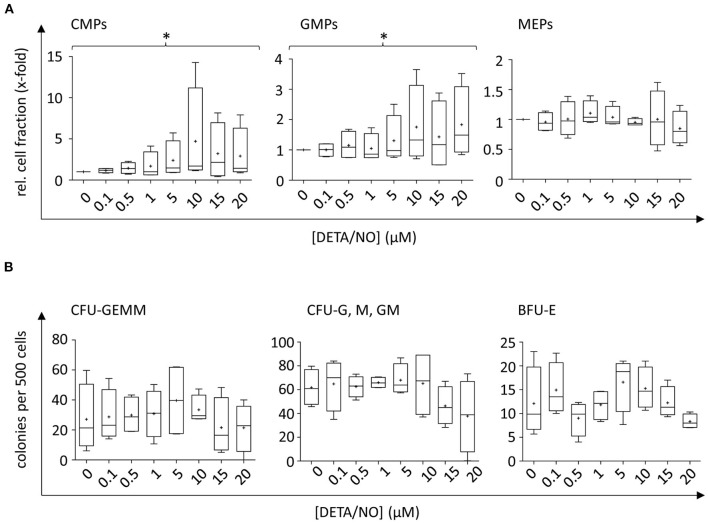
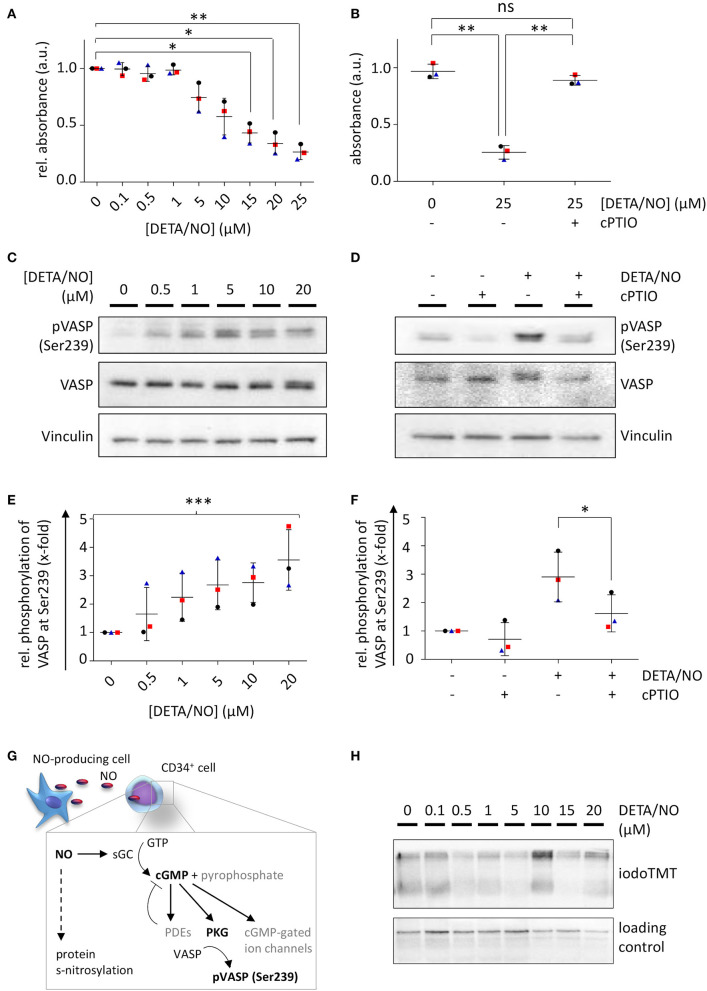
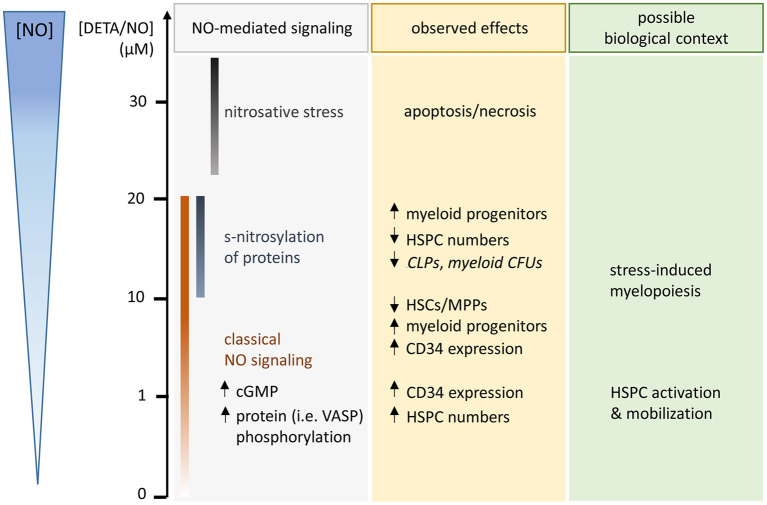
Hematopoietic stem and progenitor cell (HSPC) transplantation is the best-studied cellular therapy and successful in vitro control of HSPCs has wide clinical implications. Nitric oxide (NO) is a central signaling molecule in vivo and has been implicated in HSPC mobilization to the blood stream in mice. The influence of NO on HSPC behavior in vitro is, however, largely obscure due to the variety of employed cell types, NO administration systems, and used concentration ranges in the literature. Additionally, most studies are based on murine cells, which do not necessarily mimic human HSPC behavior. Thus, the aim of the present study was the systematic, concentration-dependent evaluation of NO-mediated effects on human HSPC behavior in vitro. By culture in the presence of the long-term NO donor diethylenetriamine/nitric oxide adduct (DETA/NO) in a nontoxic concentration window, a biphasic role of NO in the regulation of HSPC behavior was identified: Low DETA/NO concentrations activated classical NO signaling, identified via increased intracellular cyclic guanosine monophosphate (cGMP) levels and proteinkinases G (PKG)-dependent vasodilator-stimulated phosphoprotein (VASP) phosphorylation and mediated a pro-proliferative response of HSPCs. In contrast, elevated NO concentrations slowed cell proliferation and induced HSPC differentiation. At high concentrations, s-nitrosylation levels were elevated, and myeloid differentiation was increased at the expense of lymphoid progenitors. Together, these findings hint at a central role of NO in regulating human HSPC behavior and stress the importance and the potential of the use of adequate NO concentrations for in vitro cultures of HSPCs, with possible implications for clinical application of in vitro expanded or differentiated HSPCs for cellular therapies.
Keywords: ROS/RNS; differentiation; gasotransmitter; hematopoietic stem cells; nitric oxide; proliferation.
Copyright © 2021 Hümmer, Kraus, Brändle and Lee-Thedieck.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
EphA5 and EphA7 forward signaling enhances human hematopoietic stem and progenitor cell maintenance, migration, and adhesion via Rac1 activation.Exp Hematol. 2017 Apr;48:72-78. doi: 10.1016/j.exphem.2016.12.001. Epub 2016 Dec 14. Exp Hematol. 2017. PMID: 27988259
-
Acute exercise mobilizes hematopoietic stem and progenitor cells and alters the mesenchymal stromal cell secretome.J Appl Physiol (1985). 2016 Mar 15;120(6):624-32. doi: 10.1152/japplphysiol.00925.2015. Epub 2016 Jan 7. J Appl Physiol (1985). 2016. PMID: 26744505
-
Bnip3lb-driven mitophagy sustains expansion of the embryonic hematopoietic stem cell pool.bioRxiv [Preprint]. 2024 Sep 23:2024.09.23.614531. doi: 10.1101/2024.09.23.614531. bioRxiv. 2024. PMID: 39386657 Free PMC article. Preprint.
-
Hematopoietic Niche - Exploring Biomimetic Cues to Improve the Functionality of Hematopoietic Stem/Progenitor Cells.Biotechnol J. 2018 Feb;13(2). doi: 10.1002/biot.201700088. Epub 2017 Dec 28. Biotechnol J. 2018. PMID: 29178199 Review.
-
Current Developments in Mobilization of Hematopoietic Stem and Progenitor Cells and Their Interaction with Niches in Bone Marrow.Transfus Med Hemother. 2017 Jun;44(3):151-164. doi: 10.1159/000477262. Epub 2017 May 29. Transfus Med Hemother. 2017. PMID: 28626366 Free PMC article. Review.
Cited by
-
Rnf111 has a pivotal role in regulating development of definitive hematopoietic stem and progenitor cells through the Smad2/3-Gcsfr/NO axis in zebrafish.Haematologica. 2025 Feb 1;110(2):385-396. doi: 10.3324/haematol.2024.285438. Haematologica. 2025. PMID: 39363867 Free PMC article.
-
Single-cell proteo-transcriptomic profiling reveals altered characteristics of stem and progenitor cells in patients receiving cytoreductive hydroxyurea in early-phase chronic myeloid leukemia.Haematologica. 2025 Jan 1;110(1):117-128. doi: 10.3324/haematol.2024.285071. Haematologica. 2025. PMID: 39157872 Free PMC article.
-
NOS2-derived low levels of NO drive psoriasis pathogenesis.Cell Death Dis. 2024 Jun 26;15(6):449. doi: 10.1038/s41419-024-06842-z. Cell Death Dis. 2024. PMID: 38926337 Free PMC article.
-
Physical Exercise and Cardiac Repair: The Potential Role of Nitric Oxide in Boosting Stem Cell Regenerative Biology.Antioxidants (Basel). 2021 Jun 23;10(7):1002. doi: 10.3390/antiox10071002. Antioxidants (Basel). 2021. PMID: 34201562 Free PMC article. Review.
-
Differential Response and Recovery Dynamics of HSPC Populations Following Plasmodium chabaudi Infection.Int J Mol Sci. 2025 Mar 20;26(6):2816. doi: 10.3390/ijms26062816. Int J Mol Sci. 2025. PMID: 40141458 Free PMC article.
References
-
- Attar A. (2014). Changes in the cell surface markers during normal hematopoiesis: a guide to cell isolation. Global J. Hematol. Blood Trans. 1, 20–28. 10.15379/2408-9877.2014.01.01.4 - DOI
-
- Batista W. L., Ogata F. T., Curcio M. F., Miguel R. B., Arai R. J., Matsuo A. L., et al. . (2013). S-nitrosoglutathione and endothelial nitric oxide synthase-derived nitric oxide regulate compartmentalized ras S-nitrosylation and stimulate cell proliferation. Antioxid. Redox Signal. 18, 221–238. 10.1089/ars.2011.4455 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases