

Bil2 Is a Novel Inhibitor of the Yeast Formin Bnr1 Required for Proper Actin Cable Organization and Polarized Secretion
- PMID: 33634134
- PMCID: PMC7900418
- DOI: 10.3389/fcell.2021.634587
Bil2 Is a Novel Inhibitor of the Yeast Formin Bnr1 Required for Proper Actin Cable Organization and Polarized Secretion
Abstract
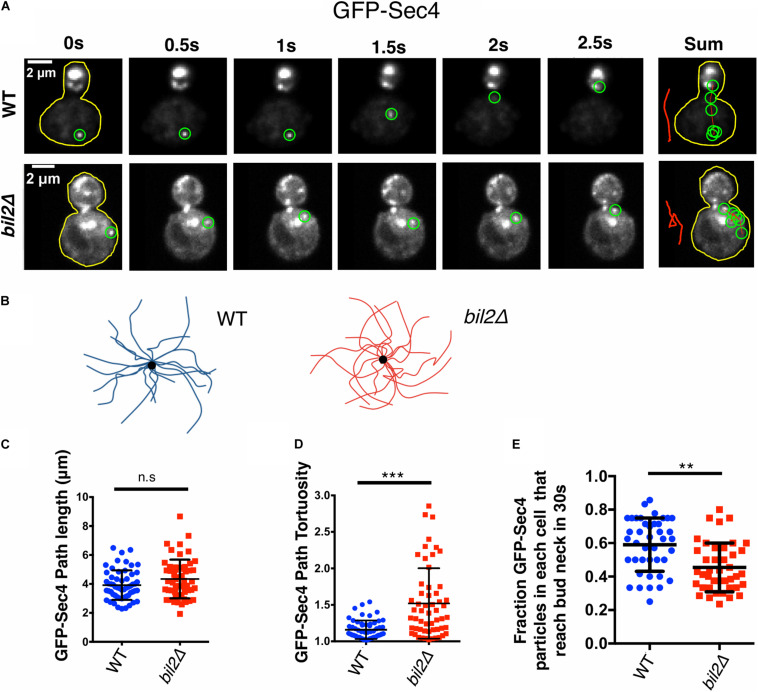
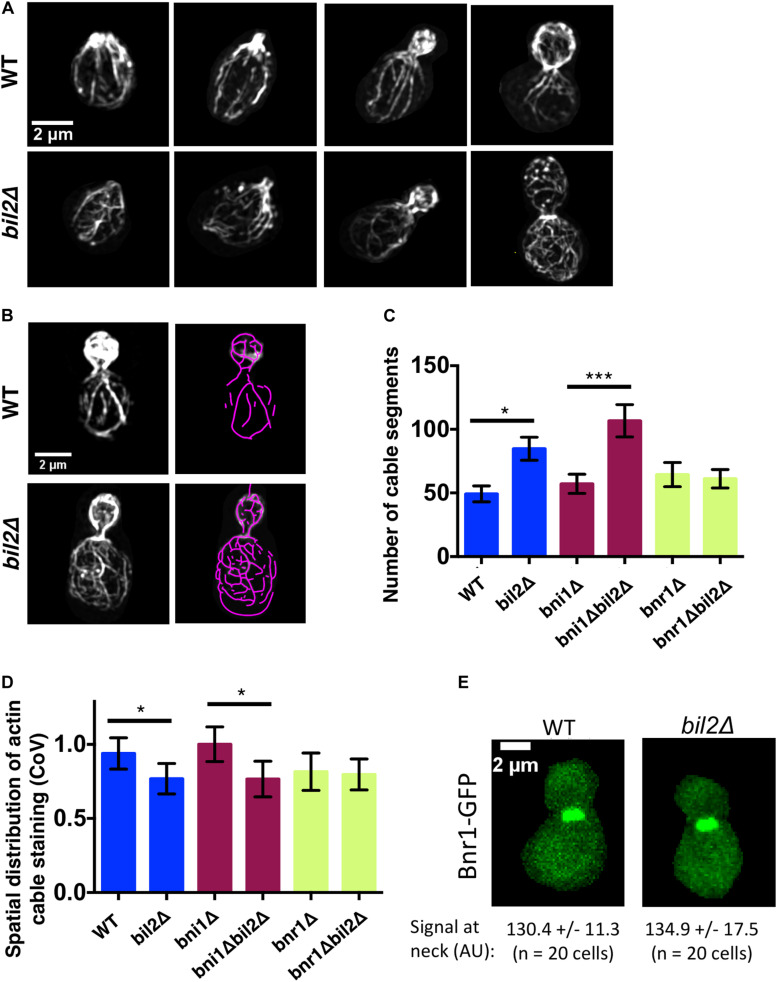
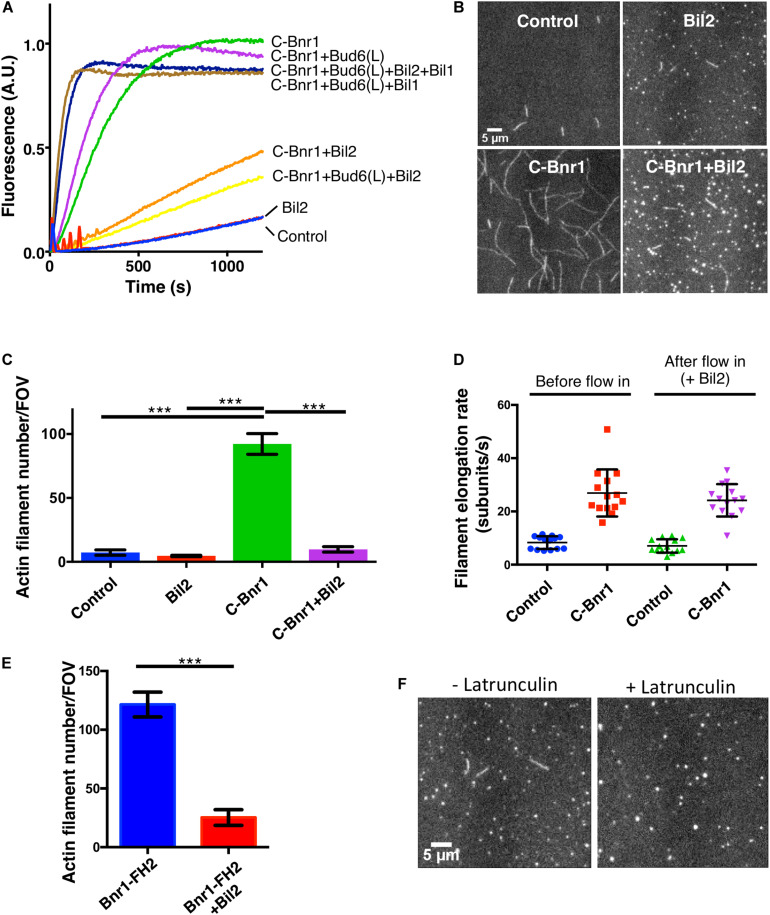
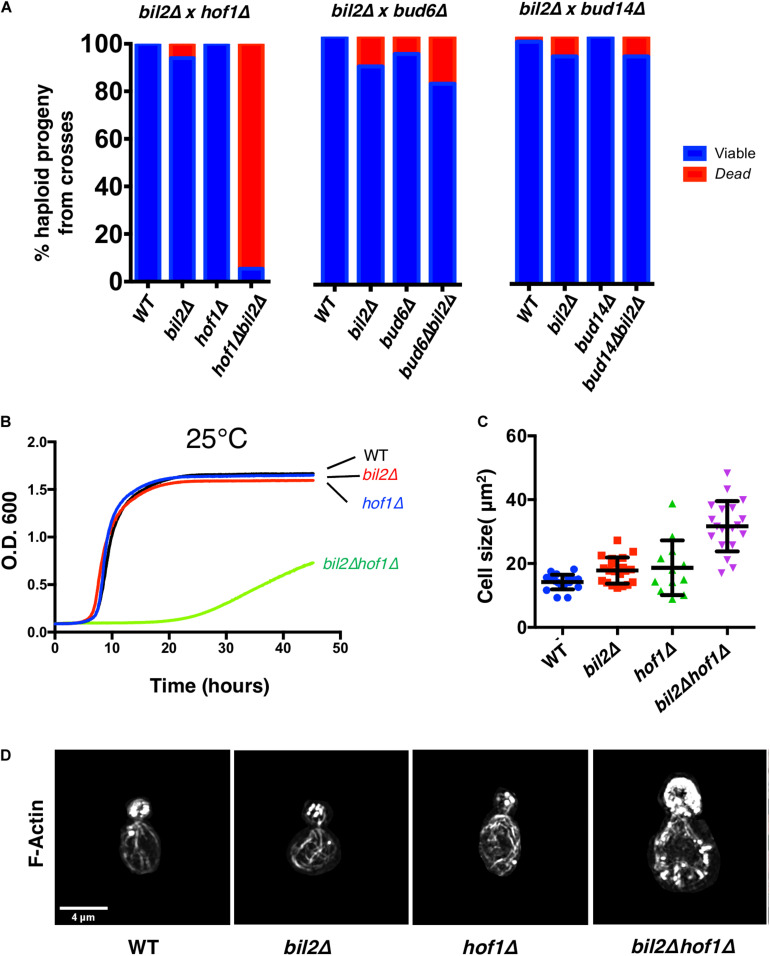
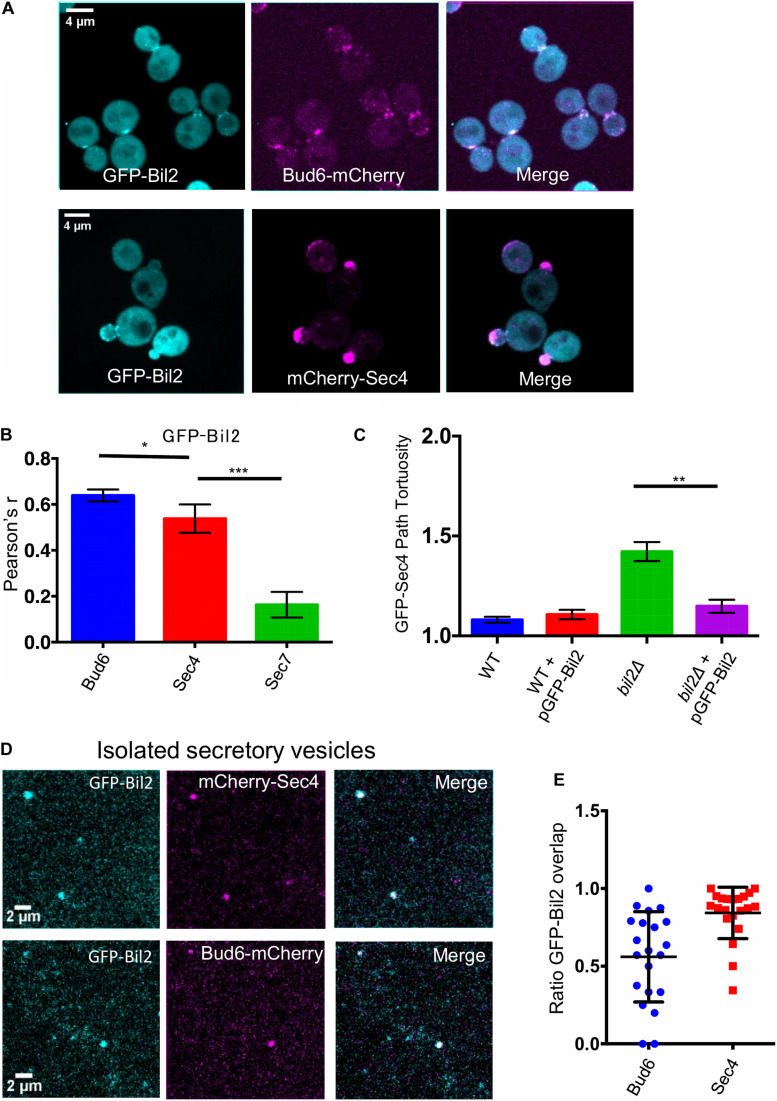
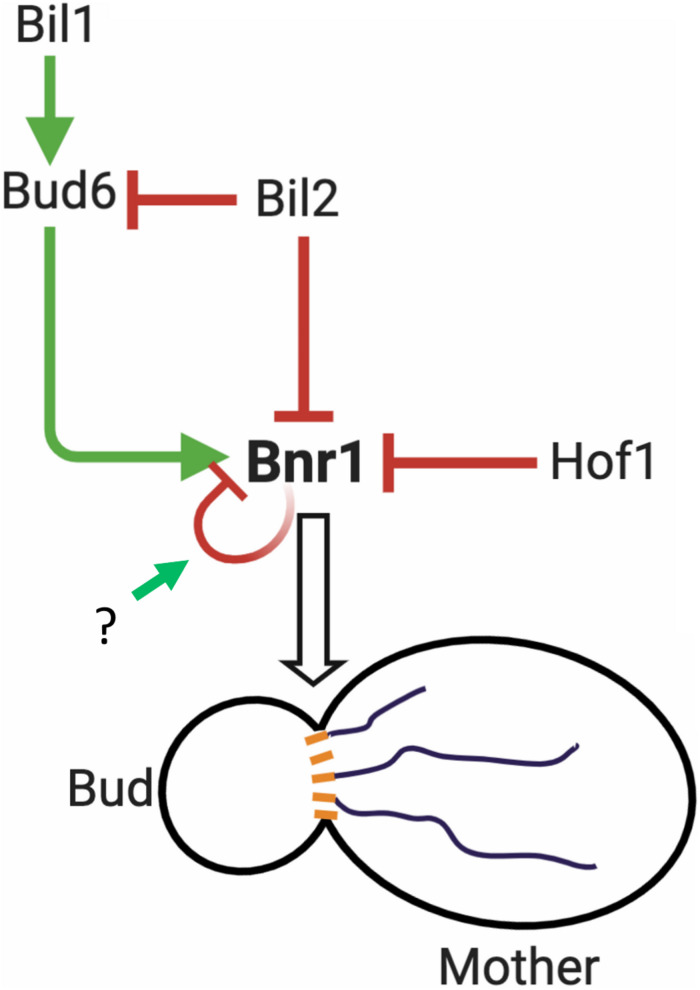
Cell growth in budding yeast depends on rapid and on-going assembly and turnover of polarized actin cables, which direct intracellular transport of post-Golgi vesicles to the bud tip. Saccharomyces cerevisiae actin cables are polymerized by two formins, Bni1 and Bnr1. Bni1 assembles cables in the bud, while Bnr1 is anchored to the bud neck and assembles cables that specifically extend filling the mother cell. Here, we report a formin regulatory role for YGL015c, a previously uncharacterized open reading frame, which we have named Bud6 Interacting Ligand 2 (BIL2). bil2Δ cells display defects in actin cable architecture and partially-impaired secretory vesicle transport. Bil2 inhibits Bnr1-mediated actin filament nucleation in vitro, yet has no effect on the rate of Bnr1-mediated filament elongation. This activity profile for Bil2 resembles that of another yeast formin regulator, the F-BAR protein Hof1, and we find that bil2Δ with hof1Δ are synthetic lethal. Unlike Hof1, which localizes exclusively to the bud neck, GFP-Bil2 localizes to the cytosol, secretory vesicles, and sites of polarized cell growth. Further, we provide evidence that Hof1 and Bil2 inhibitory effects on Bnr1 are overcome by distinct mechanisms. Together, our results suggest that Bil2 and Hof1 perform distinct yet genetically complementary roles in inhibiting the actin nucleation activity of Bnr1 to control actin cable assembly and polarized secretion.
Keywords: Bil2; Bni1; Bnr1; Bud6; actin; cable; formin; secretion.
Copyright © 2021 Rands and Goode.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The roles of yeast formins and their regulators Bud6 and Bil2 in the pheromone response.Mol Biol Cell. 2024 Jun 1;35(6):ar85. doi: 10.1091/mbc.E23-11-0459. Epub 2024 Apr 24. Mol Biol Cell. 2024. PMID: 38656798 Free PMC article.
-
Differential activities and regulation of Saccharomyces cerevisiae formin proteins Bni1 and Bnr1 by Bud6.J Biol Chem. 2005 Jul 29;280(30):28023-33. doi: 10.1074/jbc.M503094200. Epub 2005 May 27. J Biol Chem. 2005. PMID: 15923184
-
The F-BAR protein Hof1 tunes formin activity to sculpt actin cables during polarized growth.Mol Biol Cell. 2014 Jun;25(11):1730-43. doi: 10.1091/mbc.E14-03-0850. Epub 2014 Apr 9. Mol Biol Cell. 2014. PMID: 24719456 Free PMC article.
-
Polarisome assembly mediates actin remodeling during polarized yeast and fungal growth.J Cell Sci. 2021 Jan 8;134(1):jcs247916. doi: 10.1242/jcs.247916. J Cell Sci. 2021. PMID: 33419950 Review.
-
Polarized distribution of intracellular components by class V myosins in Saccharomyces cerevisiae.Int Rev Cytol. 2003;229:1-42. doi: 10.1016/s0074-7696(03)29001-x. Int Rev Cytol. 2003. PMID: 14669953 Review.
Cited by
-
Evolutionary tuning of barbed end competition allows simultaneous construction of architecturally distinct actin structures.J Cell Biol. 2023 Apr 3;222(4):e202209105. doi: 10.1083/jcb.202209105. Epub 2023 Feb 2. J Cell Biol. 2023. PMID: 36729023 Free PMC article.
-
Sequential recruitment of F-BAR proteins controls cytoskeletal crosstalk at the yeast bud neck.Curr Biol. 2025 Feb 3;35(3):574-590.e10. doi: 10.1016/j.cub.2024.12.011. Epub 2025 Jan 10. Curr Biol. 2025. PMID: 39798561
-
Nuclear protein FNBP4: A novel inhibitor of non-diaphanous formin FMN1-mediated actin cytoskeleton dynamics.J Biol Chem. 2025 Jun;301(6):108550. doi: 10.1016/j.jbc.2025.108550. Epub 2025 Apr 30. J Biol Chem. 2025. PMID: 40316024 Free PMC article.
-
The roles of yeast formins and their regulators Bud6 and Bil2 in the pheromone response.Mol Biol Cell. 2024 Jun 1;35(6):ar85. doi: 10.1091/mbc.E23-11-0459. Epub 2024 Apr 24. Mol Biol Cell. 2024. PMID: 38656798 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous