

Dynamic Microtubule Arrays in Leukocytes and Their Role in Cell Migration and Immune Synapse Formation
- PMID: 33634136
- PMCID: PMC7900162
- DOI: 10.3389/fcell.2021.635511
Dynamic Microtubule Arrays in Leukocytes and Their Role in Cell Migration and Immune Synapse Formation
Abstract
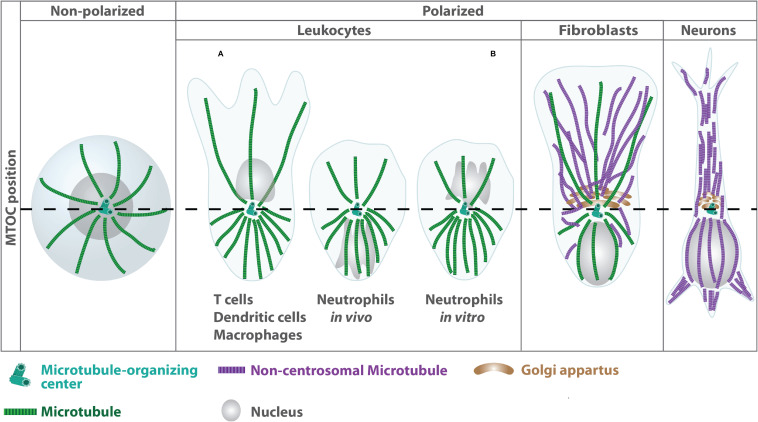
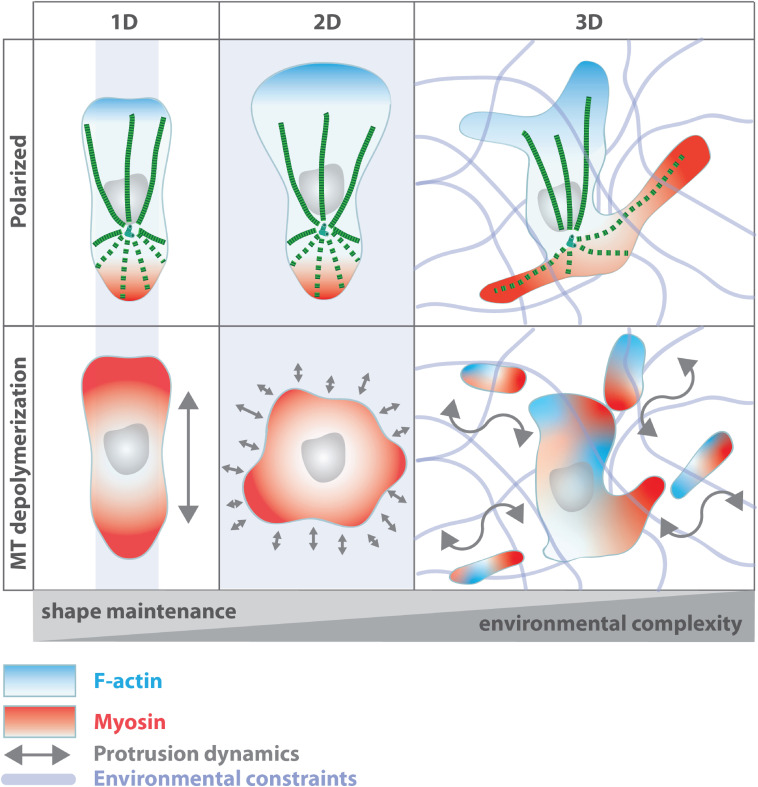
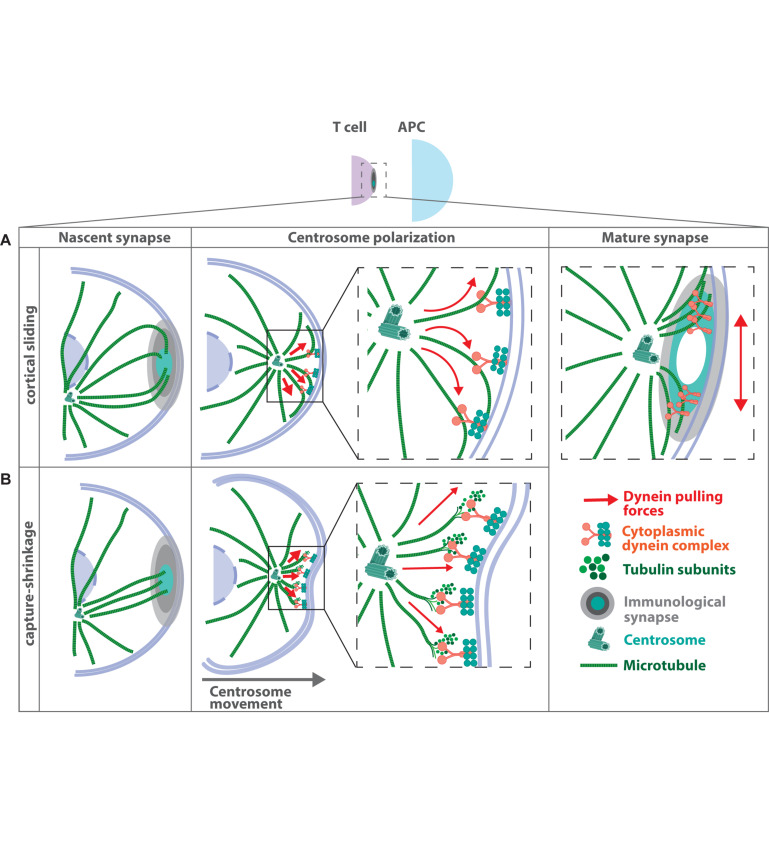
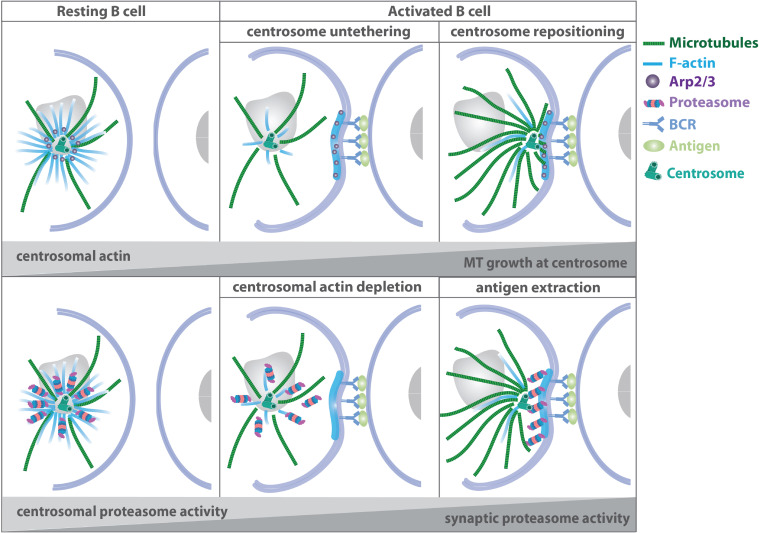
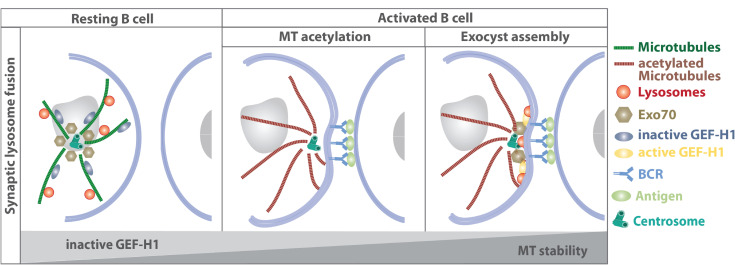
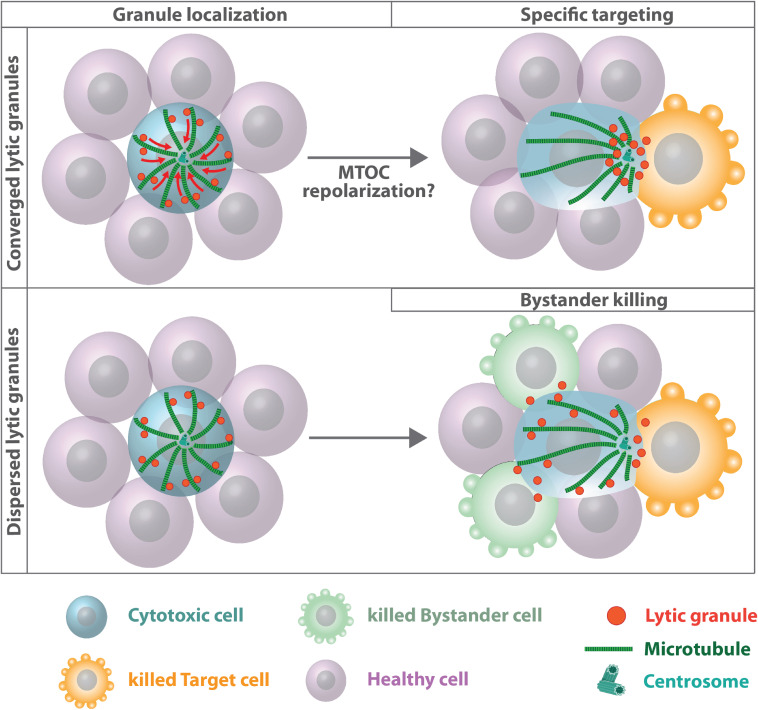
The organization of microtubule arrays in immune cells is critically important for a properly operating immune system. Leukocytes are white blood cells of hematopoietic origin, which exert effector functions of innate and adaptive immune responses. During these processes the microtubule cytoskeleton plays a crucial role for establishing cell polarization and directed migration, targeted secretion of vesicles for T cell activation and cellular cytotoxicity as well as the maintenance of cell integrity. Considering this large spectrum of distinct effector functions, leukocytes require flexible microtubule arrays, which timely and spatially reorganize allowing the cells to accommodate their specific tasks. In contrast to other specialized cell types, which typically nucleate microtubule filaments from non-centrosomal microtubule organizing centers (MTOCs), leukocytes mainly utilize centrosomes for sites of microtubule nucleation. Yet, MTOC localization as well as microtubule organization and dynamics are highly plastic in leukocytes thus allowing the cells to adapt to different environmental constraints. Here we summarize our current knowledge on microtubule organization and dynamics during immune processes and how these microtubule arrays affect immune cell effector functions. We particularly highlight emerging concepts of microtubule involvement during maintenance of cell shape and physical coherence.
Keywords: cell coherence; cell migration; immune synapse; leukocytes; microtubules.
Copyright © 2021 Kopf and Kiermaier.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Three microtubule-organizing centres collaborate in a mouse cochlear epithelial cell during supracellularly coordinated control of microtubule positioning.J Cell Sci. 1995 Jan;108 ( Pt 1):37-50. doi: 10.1242/jcs.108.1.37. J Cell Sci. 1995. PMID: 7738112
-
Microtubule-organizing centers: from the centrosome to non-centrosomal sites.Curr Opin Cell Biol. 2017 Feb;44:93-101. doi: 10.1016/j.ceb.2016.09.003. Epub 2016 Sep 22. Curr Opin Cell Biol. 2017. PMID: 27666167 Free PMC article. Review.
-
Ice recovery assay for detection of Golgi-derived microtubules.Methods Cell Biol. 2013;118:401-15. doi: 10.1016/B978-0-12-417164-0.00024-0. Methods Cell Biol. 2013. PMID: 24295320 Free PMC article.
-
Actin reorganization at the centrosomal area and the immune synapse regulates polarized secretory traffic of multivesicular bodies in T lymphocytes.J Extracell Vesicles. 2020 Jun 19;9(1):1759926. doi: 10.1080/20013078.2020.1759926. J Extracell Vesicles. 2020. PMID: 32939232 Free PMC article.
-
Centrosomal and Non-Centrosomal Microtubule-Organizing Centers (MTOCs) in Drosophila melanogaster.Cells. 2018 Aug 28;7(9):121. doi: 10.3390/cells7090121. Cells. 2018. PMID: 30154378 Free PMC article. Review.
Cited by
-
Therapeutic gene silencing of CKAP5 leads to lethality in genetically unstable cancer cells.Sci Adv. 2023 Apr 5;9(14):eade4800. doi: 10.1126/sciadv.ade4800. Epub 2023 Apr 5. Sci Adv. 2023. PMID: 37018392 Free PMC article.
-
Tubulin Post-Translational Modifications: The Elusive Roles of Acetylation.Biology (Basel). 2023 Apr 6;12(4):561. doi: 10.3390/biology12040561. Biology (Basel). 2023. PMID: 37106761 Free PMC article. Review.
-
Aging of the Immune System: Focus on Natural Killer Cells Phenotype and Functions.Cells. 2022 Mar 17;11(6):1017. doi: 10.3390/cells11061017. Cells. 2022. PMID: 35326467 Free PMC article. Review.
-
Locked and Loaded: Mechanisms Regulating Natural Killer Cell Lytic Granule Biogenesis and Release.Front Immunol. 2022 Apr 26;13:871106. doi: 10.3389/fimmu.2022.871106. eCollection 2022. Front Immunol. 2022. PMID: 35558071 Free PMC article. Review.
-
S100A8/S100A9 Integrates F-Actin and Microtubule Dynamics to Prevent Uncontrolled Extravasation of Leukocytes.Biomedicines. 2023 Mar 9;11(3):835. doi: 10.3390/biomedicines11030835. Biomedicines. 2023. PMID: 36979814 Free PMC article.
References
-
- Abercrombie M., Dunn G. A., Heath J. P. (1977). The shape and movement of fibroblasts in culture. Soc. Gen. Physiol. Ser. 32 57–70. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources