

Forkhead transcription factor Fkh1: insights into functional regulatory domains crucial for recruitment of Sin3 histone deacetylase complex
- PMID: 33635403
- PMCID: PMC8139909
- DOI: 10.1007/s00294-021-01158-3
Forkhead transcription factor Fkh1: insights into functional regulatory domains crucial for recruitment of Sin3 histone deacetylase complex
Abstract
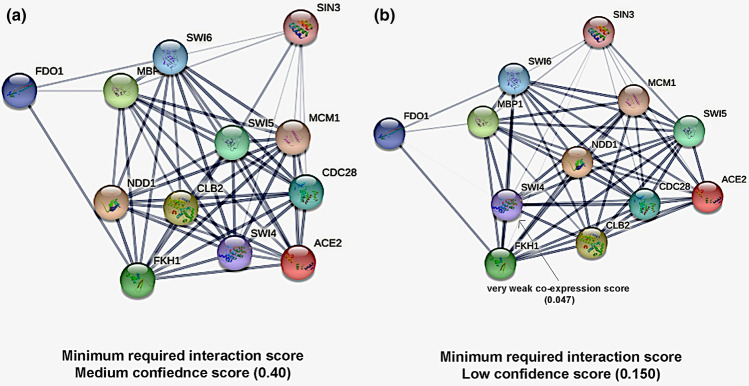
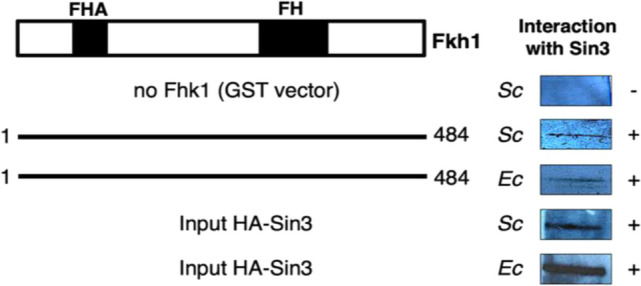
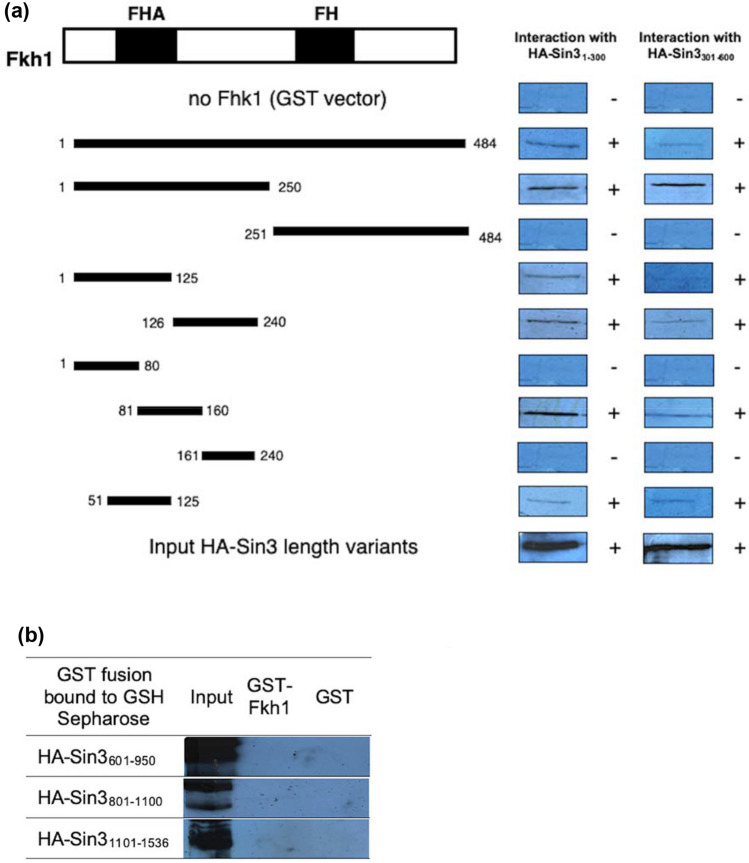
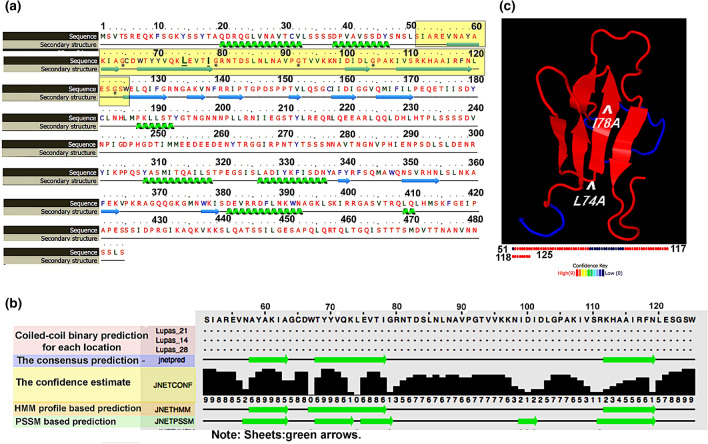
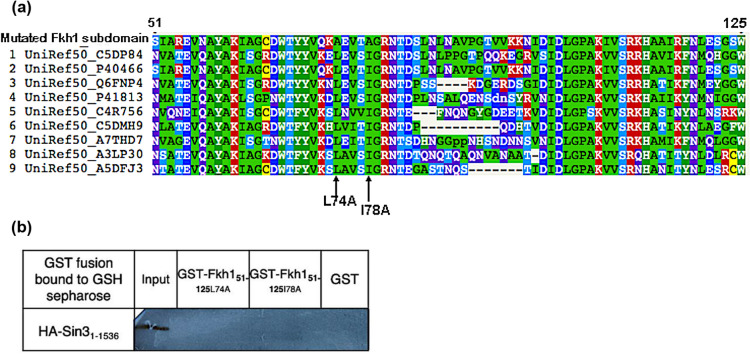
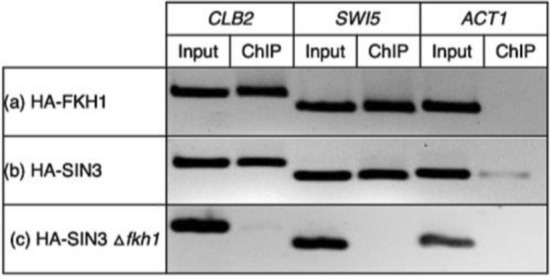
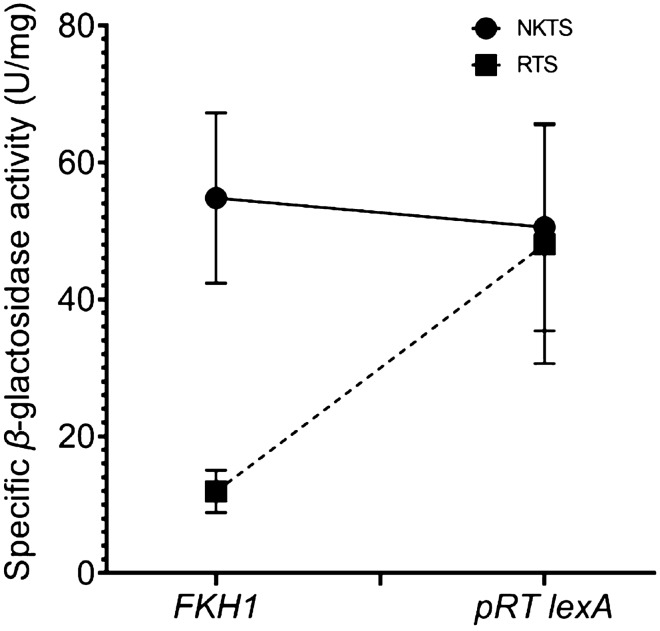
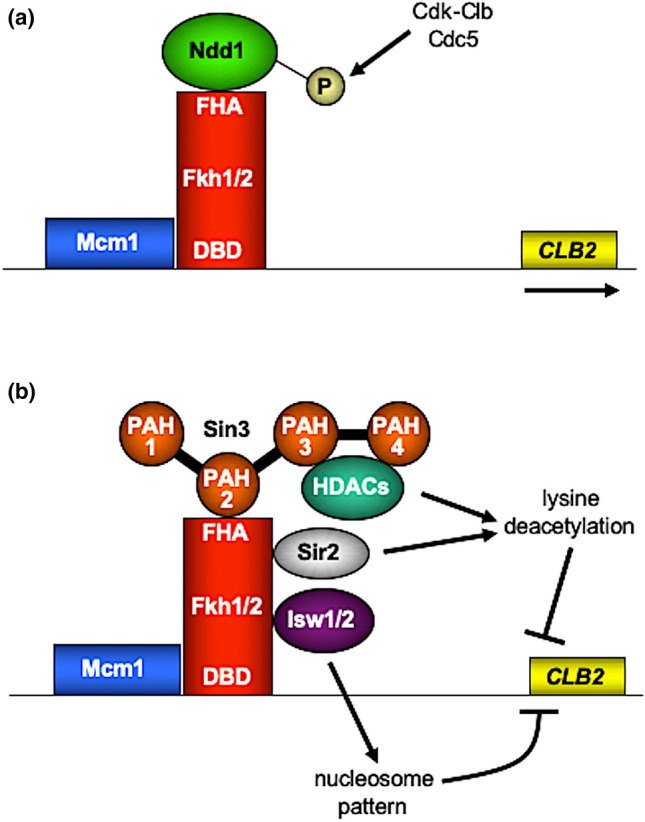
Transcription factors are inextricably linked with histone deacetylases leading to compact chromatin. The Forkhead transcription factor Fkh1 is mainly a negative transcriptional regulator which affects cell cycle control, silencing of mating-type cassettes and induction of pseudohyphal growth in the yeast Saccharomyces cerevisiae. Markedly, Fkh1 impinges chromatin architecture by recruiting large regulatory complexes. Implication of Fkh1 with transcriptional corepressor complexes remains largely unexplored. In this work we show that Fkh1 directly recruits corepressors Sin3 and Tup1 (but not Cyc8), providing evidence for its influence on epigenetic regulation. We also identified the specific domain of Fkh1 mediating Sin3 recruitment and substantiated that amino acids 51-125 of Fkh1 bind PAH2 of Sin3. Importantly, this part of Fkh1 overlaps with its Forkhead-associated domain (FHA). To analyse this domain in more detail, selected amino acids were replaced by alanine, revealing that hydrophobic amino acids L74 and I78 are important for Fkh1-Sin3 binding. In addition, we could prove Fkh1 recruitment to promoters of cell cycle genes CLB2 and SWI5. Notably, Sin3 is also recruited to these promoters but only in the presence of functional Fkh1. Our results disclose that recruitment of Sin3 to Fkh1 requires precisely positioned Fkh1/Sin3 binding sites which provide an extended view on the genetic control of cell cycle genes CLB2 and SWI5 and the mechanism of transcriptional repression by modulation of chromatin architecture at the G2/M transition.
Keywords: Cell cycle genes; Fkh1; Histone deacetylases (HDACs); Sin3; Tup1.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
