

Sustained neural rhythms reveal endogenous oscillations supporting speech perception
- PMID: 33635855
- PMCID: PMC7946281
- DOI: 10.1371/journal.pbio.3001142
Sustained neural rhythms reveal endogenous oscillations supporting speech perception
Abstract
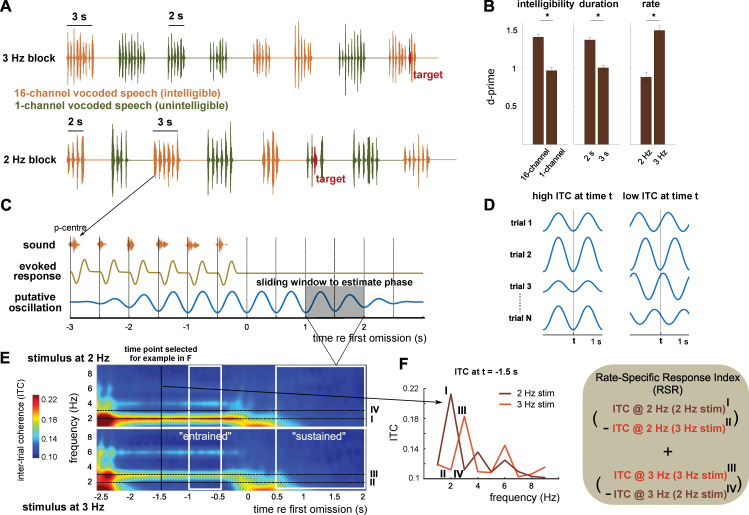
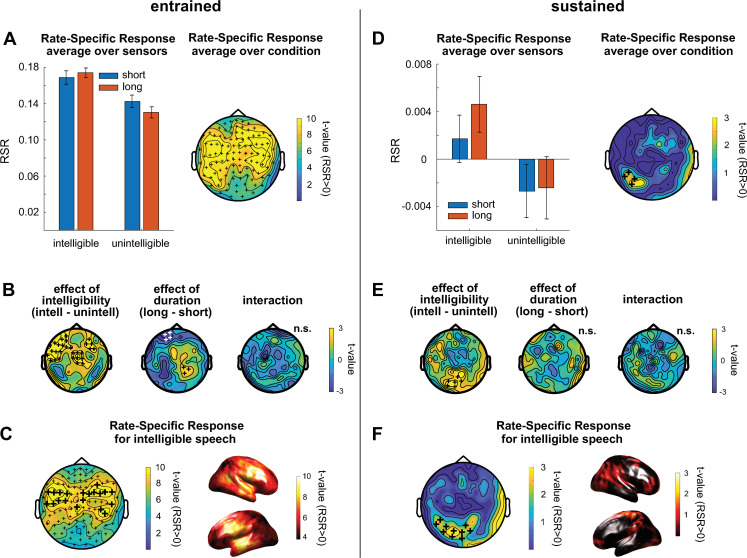
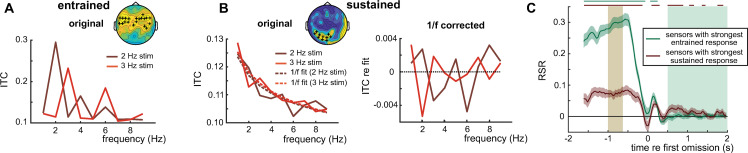
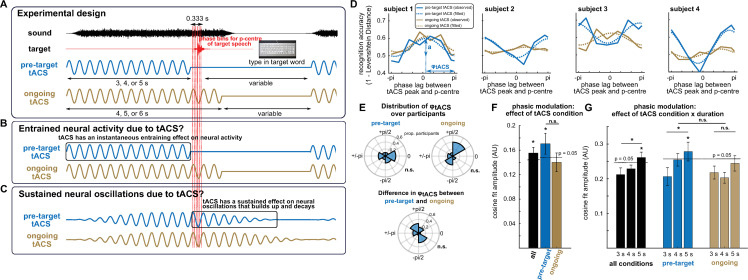
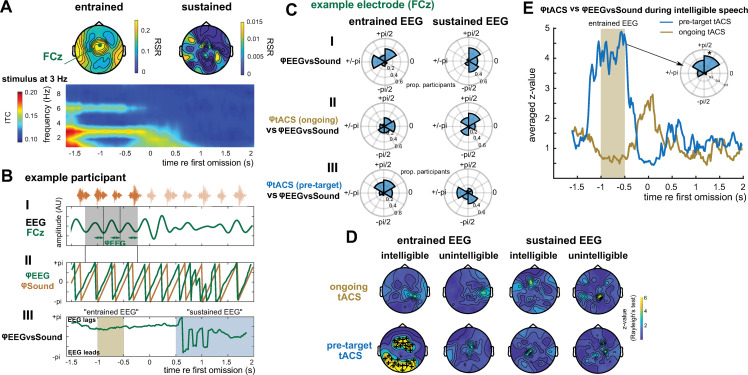
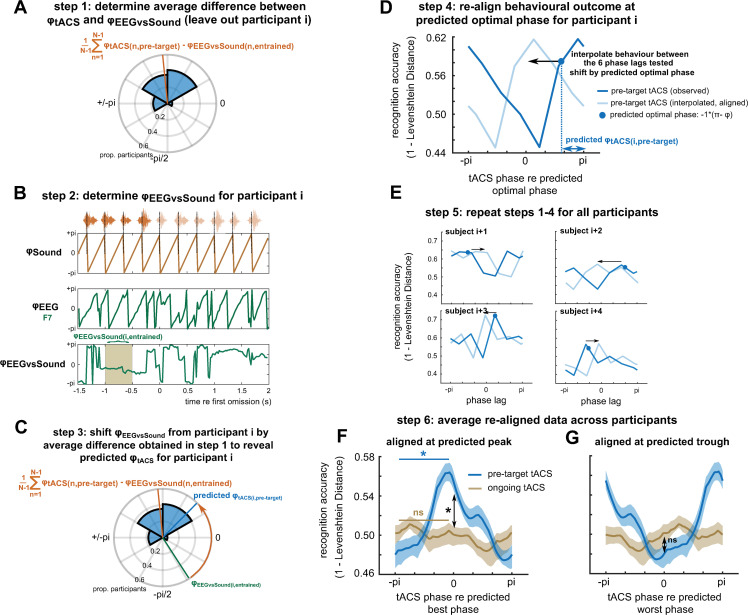
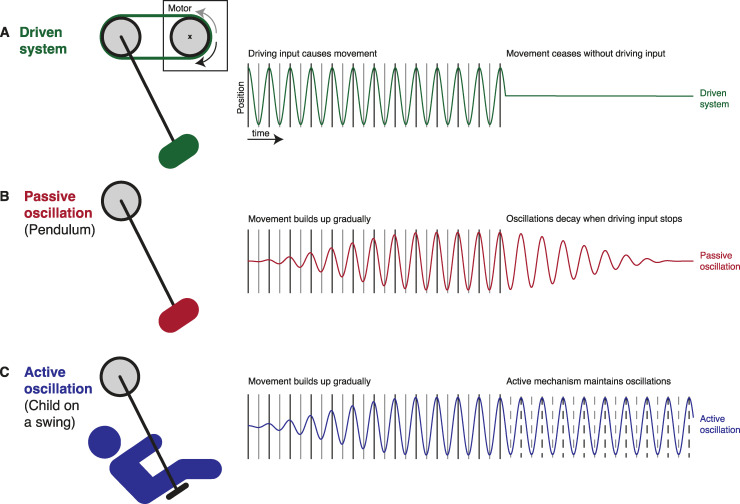
Rhythmic sensory or electrical stimulation will produce rhythmic brain responses. These rhythmic responses are often interpreted as endogenous neural oscillations aligned (or "entrained") to the stimulus rhythm. However, stimulus-aligned brain responses can also be explained as a sequence of evoked responses, which only appear regular due to the rhythmicity of the stimulus, without necessarily involving underlying neural oscillations. To distinguish evoked responses from true oscillatory activity, we tested whether rhythmic stimulation produces oscillatory responses which continue after the end of the stimulus. Such sustained effects provide evidence for true involvement of neural oscillations. In Experiment 1, we found that rhythmic intelligible, but not unintelligible speech produces oscillatory responses in magnetoencephalography (MEG) which outlast the stimulus at parietal sensors. In Experiment 2, we found that transcranial alternating current stimulation (tACS) leads to rhythmic fluctuations in speech perception outcomes after the end of electrical stimulation. We further report that the phase relation between electroencephalography (EEG) responses and rhythmic intelligible speech can predict the tACS phase that leads to most accurate speech perception. Together, we provide fundamental results for several lines of research-including neural entrainment and tACS-and reveal endogenous neural oscillations as a key underlying principle for speech perception.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Phase Entrainment of Brain Oscillations Causally Modulates Neural Responses to Intelligible Speech.Curr Biol. 2018 Feb 5;28(3):401-408.e5. doi: 10.1016/j.cub.2017.11.071. Epub 2018 Jan 18. Curr Biol. 2018. PMID: 29358073 Free PMC article.
-
Friends, not foes: Magnetoencephalography as a tool to uncover brain dynamics during transcranial alternating current stimulation.Neuroimage. 2015 Sep;118:406-13. doi: 10.1016/j.neuroimage.2015.06.026. Epub 2015 Jun 12. Neuroimage. 2015. PMID: 26080310 Free PMC article.
-
Entrainment echoes in the cerebellum.Proc Natl Acad Sci U S A. 2024 Aug 20;121(34):e2411167121. doi: 10.1073/pnas.2411167121. Epub 2024 Aug 13. Proc Natl Acad Sci U S A. 2024. PMID: 39136991 Free PMC article.
-
EEG oscillations: From correlation to causality.Int J Psychophysiol. 2016 May;103:12-21. doi: 10.1016/j.ijpsycho.2015.02.003. Epub 2015 Feb 4. Int J Psychophysiol. 2016. PMID: 25659527 Review.
-
The Involvement of Endogenous Neural Oscillations in the Processing of Rhythmic Input: More Than a Regular Repetition of Evoked Neural Responses.Front Neurosci. 2018 Mar 7;12:95. doi: 10.3389/fnins.2018.00095. eCollection 2018. Front Neurosci. 2018. PMID: 29563860 Free PMC article. Review.
Cited by
-
Forward entrainment: Psychophysics, neural correlates, and function.Psychon Bull Rev. 2023 Jun;30(3):803-821. doi: 10.3758/s13423-022-02220-y. Epub 2022 Dec 2. Psychon Bull Rev. 2023. PMID: 36460893 Free PMC article. Review.
-
Opposing neural processing modes alternate rhythmically during sustained auditory attention.Commun Biol. 2024 Sep 12;7(1):1125. doi: 10.1038/s42003-024-06834-x. Commun Biol. 2024. PMID: 39266696 Free PMC article.
-
MEG Activity in Visual and Auditory Cortices Represents Acoustic Speech-Related Information during Silent Lip Reading.eNeuro. 2022 Jun 27;9(3):ENEURO.0209-22.2022. doi: 10.1523/ENEURO.0209-22.2022. Print 2022 May-Jun. eNeuro. 2022. PMID: 35728955 Free PMC article.
-
Distracting linguistic information impairs neural tracking of attended speech.Curr Res Neurobiol. 2022 May 28;3:100043. doi: 10.1016/j.crneur.2022.100043. eCollection 2022. Curr Res Neurobiol. 2022. PMID: 36518343 Free PMC article.
-
Speech Prosody Serves Temporal Prediction of Language via Contextual Entrainment.J Neurosci. 2024 Jul 10;44(28):e1041232024. doi: 10.1523/JNEUROSCI.1041-23.2024. J Neurosci. 2024. PMID: 38839302 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
