

Mendelian pathway analysis of laboratory traits reveals distinct roles for ciliary subcompartments in common disease pathogenesis
- PMID: 33636100
- PMCID: PMC8008498
- DOI: 10.1016/j.ajhg.2021.02.008
Mendelian pathway analysis of laboratory traits reveals distinct roles for ciliary subcompartments in common disease pathogenesis
Abstract
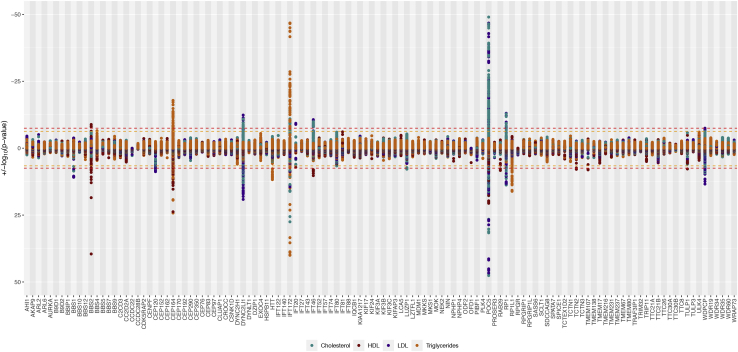
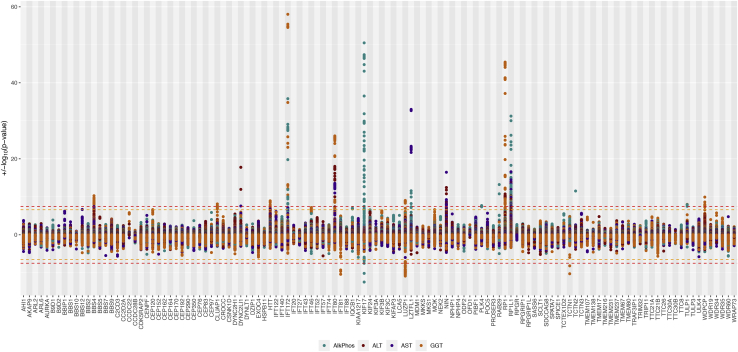
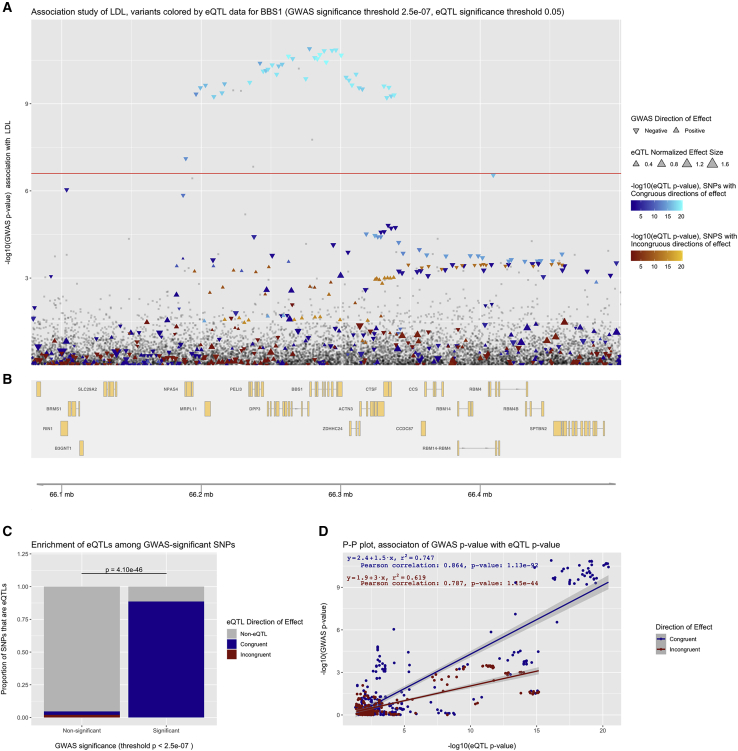
Rare monogenic disorders of the primary cilium, termed ciliopathies, are characterized by extreme presentations of otherwise common diseases, such as diabetes, hepatic fibrosis, and kidney failure. However, despite a recent revolution in our understanding of the cilium's role in rare disease pathogenesis, the organelle's contribution to common disease remains largely unknown. Hypothesizing that common genetic variants within Mendelian ciliopathy genes might contribute to common complex diseases pathogenesis, we performed association studies of 16,874 common genetic variants across 122 ciliary genes with 12 quantitative laboratory traits characteristic of ciliopathy syndromes in 452,593 individuals in the UK Biobank. We incorporated tissue-specific gene expression analysis, expression quantitative trait loci, and Mendelian disease phenotype information into our analysis and replicated our findings in meta-analysis. 101 statistically significant associations were identified across 42 of the 122 examined ciliary genes (including eight novel replicating associations). These ciliary genes were widely expressed in tissues relevant to the phenotypes being studied, and eQTL analysis revealed strong evidence for correlation between ciliary gene expression levels and laboratory traits. Perhaps most interestingly, our analysis identified different ciliary subcompartments as being specifically associated with distinct sets of phenotypes. Taken together, our data demonstrate the utility of a Mendelian pathway-based approach to genomic association studies, challenge the widely held belief that the cilium is an organelle important mainly in development and in rare syndromic disease pathogenesis, and provide a framework for the continued integration of common and rare disease genetics to provide insight into the pathophysiology of human diseases of immense public health burden.
Keywords: Mendelian disease; bioinformatics; ciliopathies; cilium; complex trait; genomics.
Copyright © 2021 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
M.D.R. is on the Scientific Advisory Board for Cipherome and for Goldfinch Bio. X.Z. is currently employed by Regeneron Pharmaceuticals, but all work for this manuscript was completed prior to her beginning in this position. The authors declare no additional competing interests.
Figures





Similar articles
-
Characterizing the morbid genome of ciliopathies.Genome Biol. 2016 Nov 28;17(1):242. doi: 10.1186/s13059-016-1099-5. Genome Biol. 2016. PMID: 27894351 Free PMC article.
-
Molecular genetics of renal ciliopathies.Biochem Soc Trans. 2021 Jun 30;49(3):1205-1220. doi: 10.1042/BST20200791. Biochem Soc Trans. 2021. PMID: 33960378 Review.
-
Insights into Ciliary Genes and Evolution from Multi-Level Phylogenetic Profiling.Mol Biol Evol. 2017 Aug 1;34(8):2016-2034. doi: 10.1093/molbev/msx146. Mol Biol Evol. 2017. PMID: 28460059 Free PMC article.
-
The Joubert-Meckel-Nephronophthisis Spectrum of Ciliopathies.Annu Rev Genomics Hum Genet. 2022 Aug 31;23:301-329. doi: 10.1146/annurev-genom-121321-093528. Epub 2022 Jun 2. Annu Rev Genomics Hum Genet. 2022. PMID: 35655331 Free PMC article. Review.
-
Mutations in ARMC9, which Encodes a Basal Body Protein, Cause Joubert Syndrome in Humans and Ciliopathy Phenotypes in Zebrafish.Am J Hum Genet. 2017 Jul 6;101(1):23-36. doi: 10.1016/j.ajhg.2017.05.010. Epub 2017 Jun 15. Am J Hum Genet. 2017. PMID: 28625504 Free PMC article.
Cited by
-
A Role for Genetic Modifiers in Tubulointerstitial Kidney Diseases.Genes (Basel). 2023 Aug 3;14(8):1582. doi: 10.3390/genes14081582. Genes (Basel). 2023. PMID: 37628633 Free PMC article. Review.
-
Keeping pace: the primary cilium as the conducting baton of the islet.Diabetologia. 2024 May;67(5):773-782. doi: 10.1007/s00125-024-06096-6. Epub 2024 Feb 14. Diabetologia. 2024. PMID: 38353726 Free PMC article. Review.
-
The Penn Medicine BioBank: Towards a Genomics-Enabled Learning Healthcare System to Accelerate Precision Medicine in a Diverse Population.J Pers Med. 2022 Nov 29;12(12):1974. doi: 10.3390/jpm12121974. J Pers Med. 2022. PMID: 36556195 Free PMC article.
-
eQTpLot: a user-friendly R package for the visualization of colocalization between eQTL and GWAS signals.BioData Min. 2021 Jul 17;14(1):32. doi: 10.1186/s13040-021-00267-6. BioData Min. 2021. PMID: 34273980 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
