

In vivo CD8+ T cell CRISPR screening reveals control by Fli1 in infection and cancer
- PMID: 33636129
- PMCID: PMC8054351
- DOI: 10.1016/j.cell.2021.02.019
In vivo CD8+ T cell CRISPR screening reveals control by Fli1 in infection and cancer
Abstract
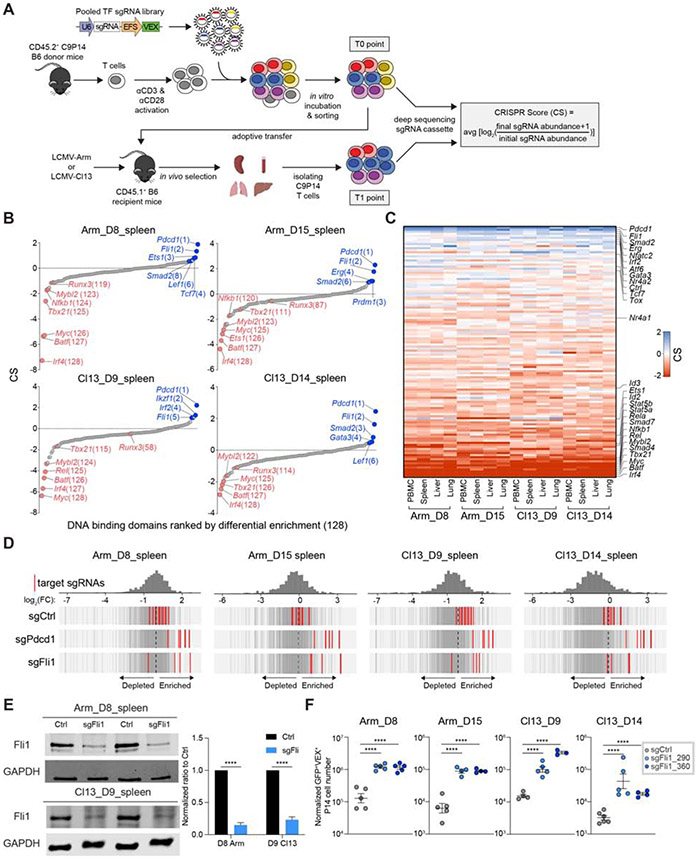
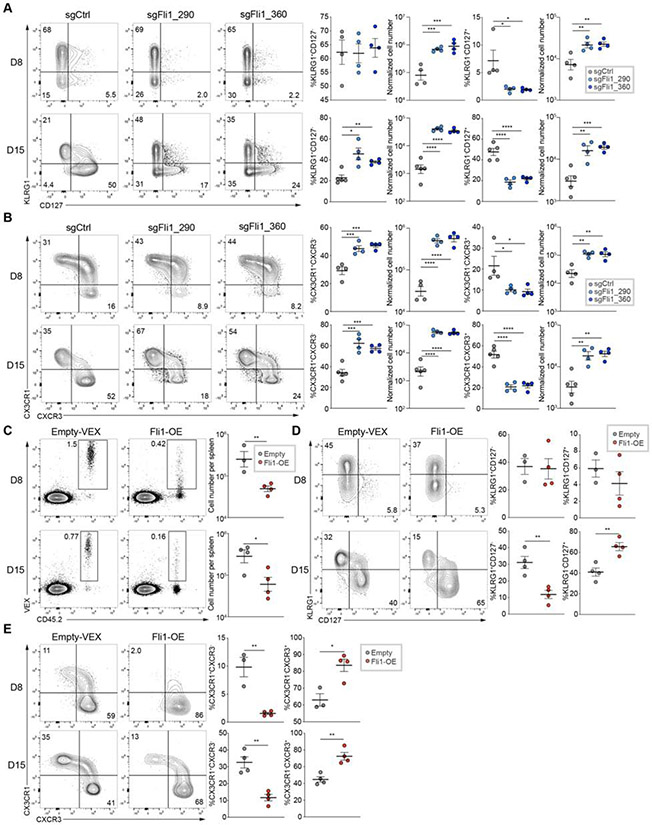
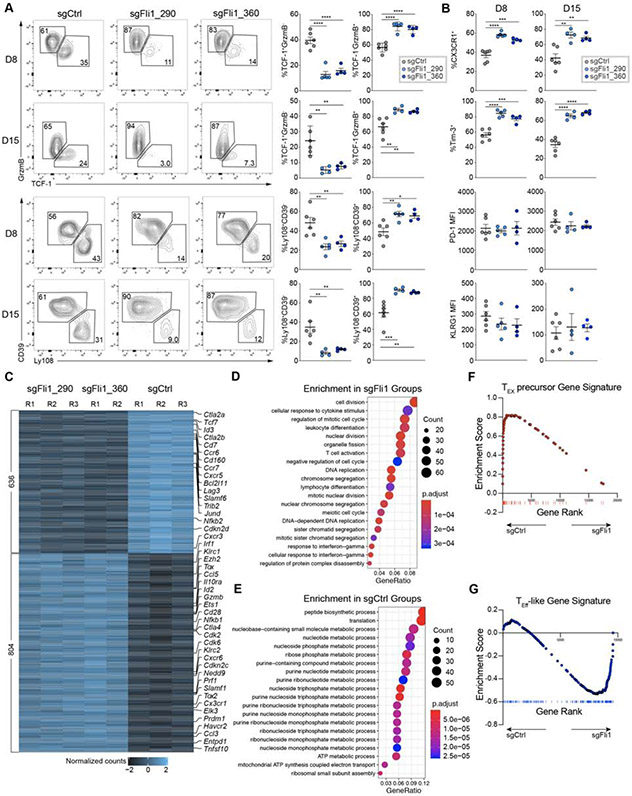
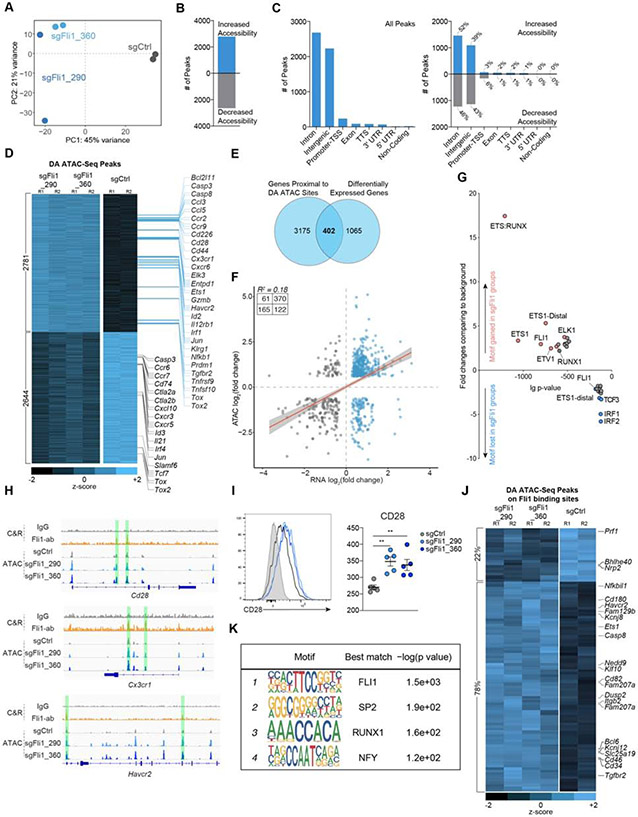
Improving effector activity of antigen-specific T cells is a major goal in cancer immunotherapy. Despite the identification of several effector T cell (TEFF)-driving transcription factors (TFs), the transcriptional coordination of TEFF biology remains poorly understood. We developed an in vivo T cell CRISPR screening platform and identified a key mechanism restraining TEFF biology through the ETS family TF, Fli1. Genetic deletion of Fli1 enhanced TEFF responses without compromising memory or exhaustion precursors. Fli1 restrained TEFF lineage differentiation by binding to cis-regulatory elements of effector-associated genes. Loss of Fli1 increased chromatin accessibility at ETS:RUNX motifs, allowing more efficient Runx3-driven TEFF biology. CD8+ T cells lacking Fli1 provided substantially better protection against multiple infections and tumors. These data indicate that Fli1 safeguards the developing CD8+ T cell transcriptional landscape from excessive ETS:RUNX-driven TEFF cell differentiation. Moreover, genetic deletion of Fli1 improves TEFF differentiation and protective immunity in infections and cancer.
Keywords: CD8; CRISPR; Fli1; cancer; chronic infection; effector CD8 T cell; exhausted CD8 T cell; exhaustion; immunotherapy; protective immunity.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.J.W. has consulting agreements with and/or is on the scientific advisory board for Merck, Elstar, Janssen, Related Sciences, Synthekine and Surface Oncology. E.J.W. is a founder of Surface Oncology and Arsenal Biosciences. E.J.W. has a patent licensing agreement on the PD-1 pathway with Roche/Genentech. O.K. is an employee and shareholder of Arsenal Biosciences.
Figures
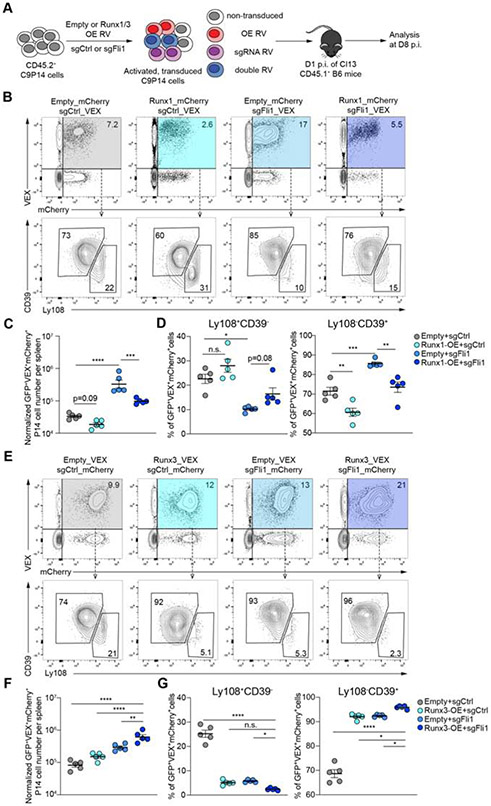
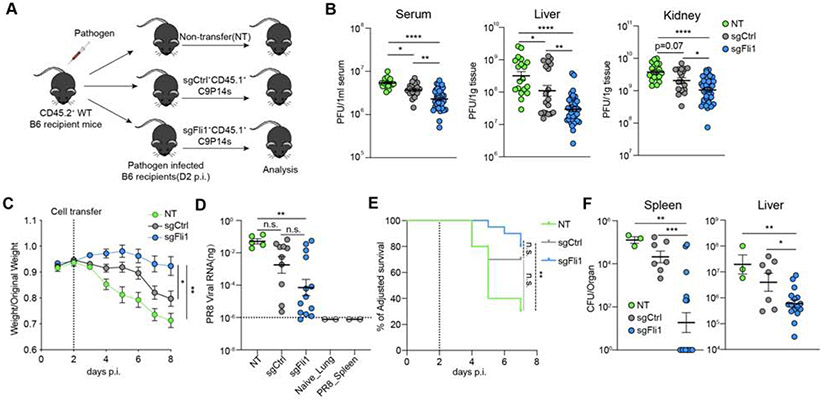
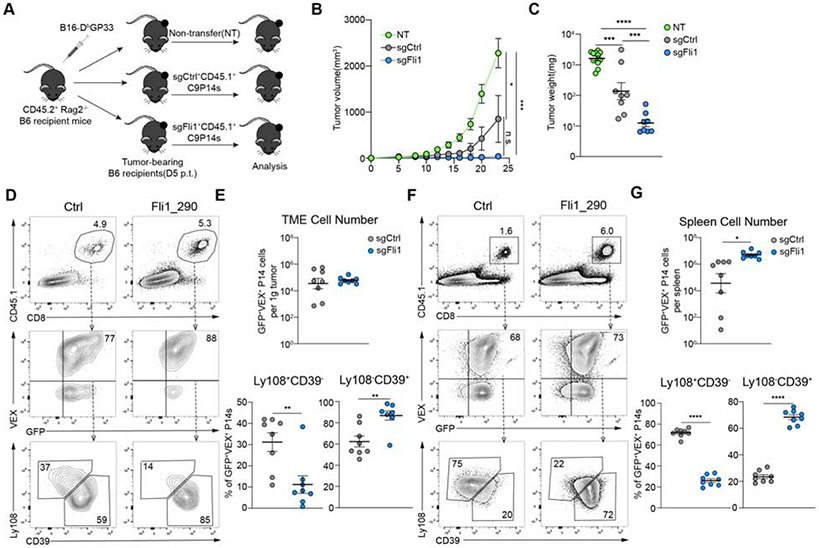
Comment in
-
How to make a better T cell: in vivo CRISPR screens have some answers.Cell. 2021 Mar 4;184(5):1135-1136. doi: 10.1016/j.cell.2021.02.003. Cell. 2021. PMID: 33667366
References
-
- Alfei F, Kanev K, Hofmann M, Wu M, Ghoneim HE, Roelli P, Utzschneider DT, Hösslin, von M, Cullen JG, Fan Y, et al. (2019). TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 2017 545:7652 1. - PubMed
-
- Badovinac VP, Porter BB, and Harty JT (2004). CD8+ T cell contraction is controlled by early inflammation. Nature Immunology 2006 7:12 5, 809–817. - PubMed
-
- Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, and Ahmed R (2006). Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 2017 545:7652 439, 682–687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 CA210944/CA/NCI NIH HHS/United States
- R01 HG010480/HG/NHGRI NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- P01 AI112521/AI/NIAID NIH HHS/United States
- R01 AI115712/AI/NIAID NIH HHS/United States
- K00 CA234842/CA/NCI NIH HHS/United States
- P01 AI108545/AI/NIAID NIH HHS/United States
- R01 AI105343/AI/NIAID NIH HHS/United States
- R01 CA078831/CA/NCI NIH HHS/United States
- R01 MH109905/MH/NIMH NIH HHS/United States
- U19 AI117950/AI/NIAID NIH HHS/United States
- T32 CA009140/CA/NCI NIH HHS/United States
- P30 AG073104/AG/NIA NIH HHS/United States
- R01 AI155577/AI/NIAID NIH HHS/United States
- F99 CA234842/CA/NCI NIH HHS/United States
- U19 AI149680/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
