

Tumor-specific cytolytic CD4 T cells mediate immunity against human cancer
- PMID: 33637530
- PMCID: PMC7909889
- DOI: 10.1126/sciadv.abe3348
Tumor-specific cytolytic CD4 T cells mediate immunity against human cancer
Abstract
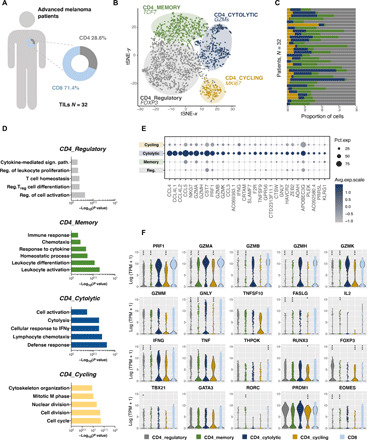
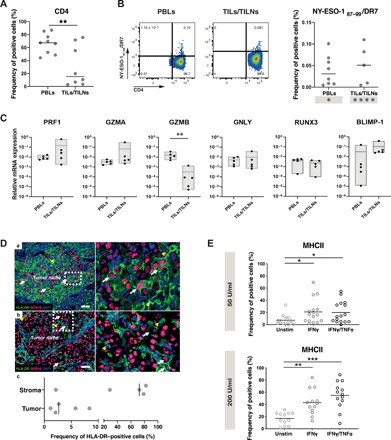
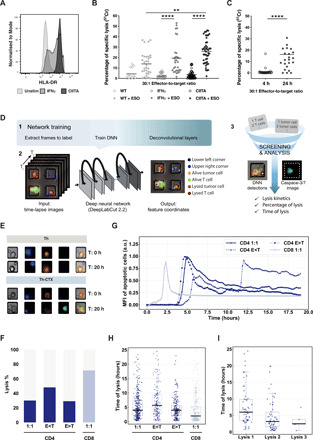
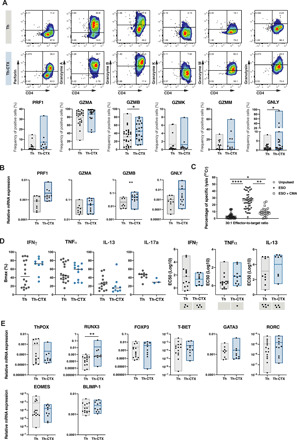
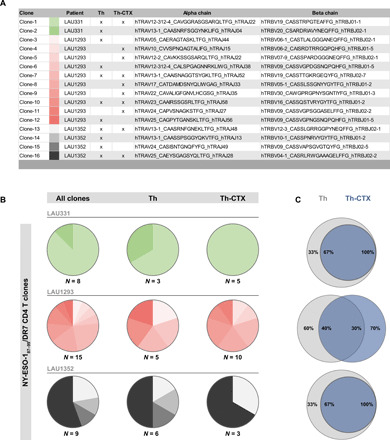
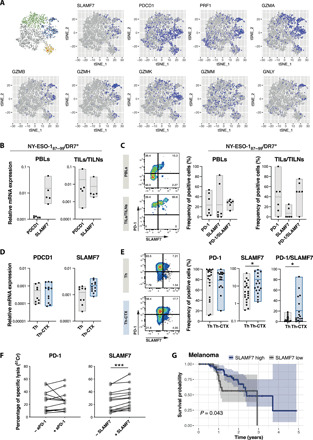
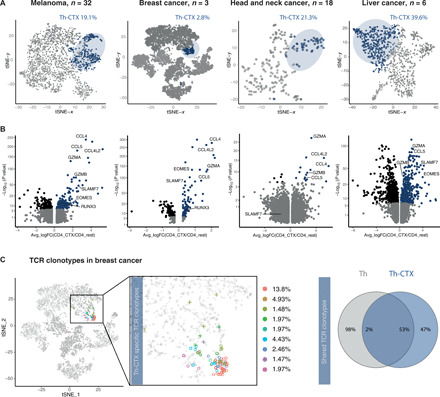
CD4 T cells have been implicated in cancer immunity for their helper functions. Moreover, their direct cytotoxic potential has been shown in some patients with cancer. Here, by mining single-cell RNA-seq datasets, we identified CD4 T cell clusters displaying cytotoxic phenotypes in different human cancers, resembling CD8 T cell profiles. Using the peptide-MHCII-multimer technology, we confirmed ex vivo the presence of cytolytic tumor-specific CD4 T cells. We performed an integrated phenotypic and functional characterization of these cells, down to the single-cell level, through a high-throughput nanobiochip consisting of massive arrays of picowells and machine learning. We demonstrated a direct, contact-, and granzyme-dependent cytotoxic activity against tumors, with delayed kinetics compared to classical cytotoxic lymphocytes. Last, we found that this cytotoxic activity was in part dependent on SLAMF7. Agonistic engagement of SLAMF7 enhanced cytotoxicity of tumor-specific CD4 T cells, suggesting that targeting these cells might prove synergistic with other cancer immunotherapies.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Sallusto F., Lanzavecchia A., Heterogeneity of CD4+ memory T cells: Functional modules for tailored immunity. Eur. J. Immunol. 39, 2076–2082 (2009). - PubMed
-
- Tran E., Turcotte S., Gros A., Robbins P. F., Lu Y. C., Dudley M. E., Wunderlich J. R., Somerville R. P., Hogan K., Hinrichs C. S., Parkhurst M. R., Yang J. C., Rosenberg S. A., Cancer immunotherapy based on mutation-specific CD4+ T cells in a patient with epithelial cancer. Science 344, 641–645 (2014). - PMC - PubMed
-
- Fridman W. H., Pagès F., Sautès-Fridman C., Galon J., The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 12, 298–306 (2012). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
