

Sema7A is crucial for resolution of severe inflammation
- PMID: 33637648
- PMCID: PMC7936270
- DOI: 10.1073/pnas.2017527118
Sema7A is crucial for resolution of severe inflammation
Abstract
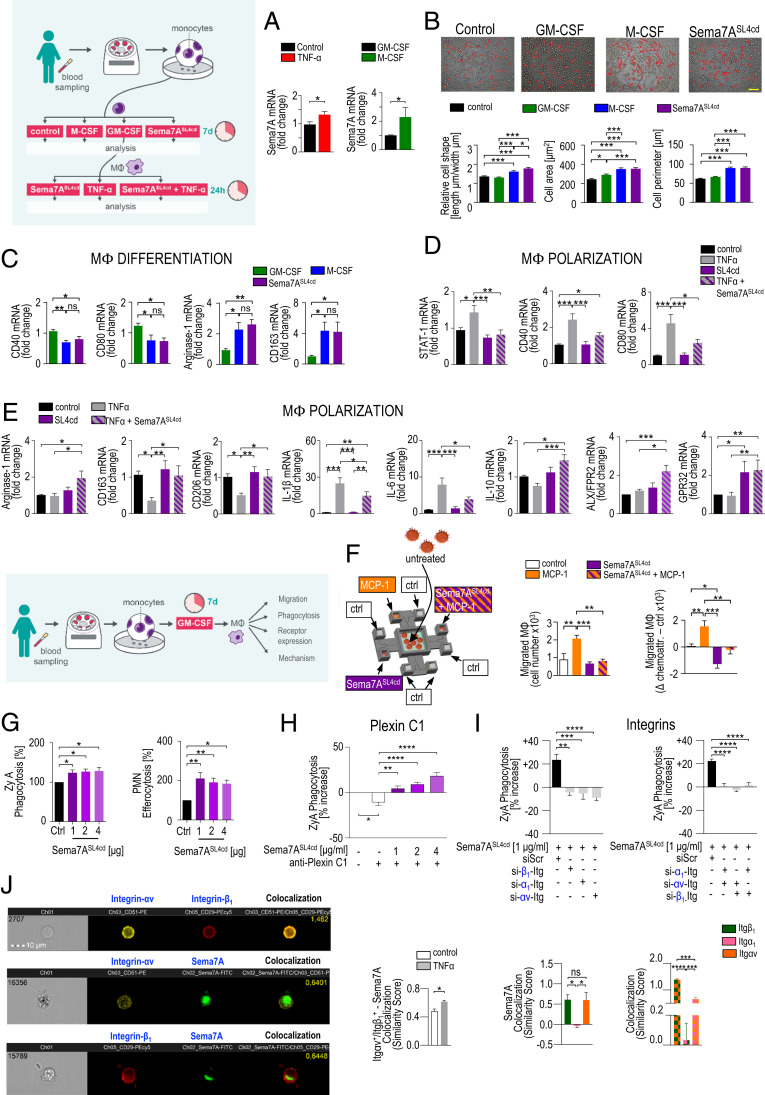
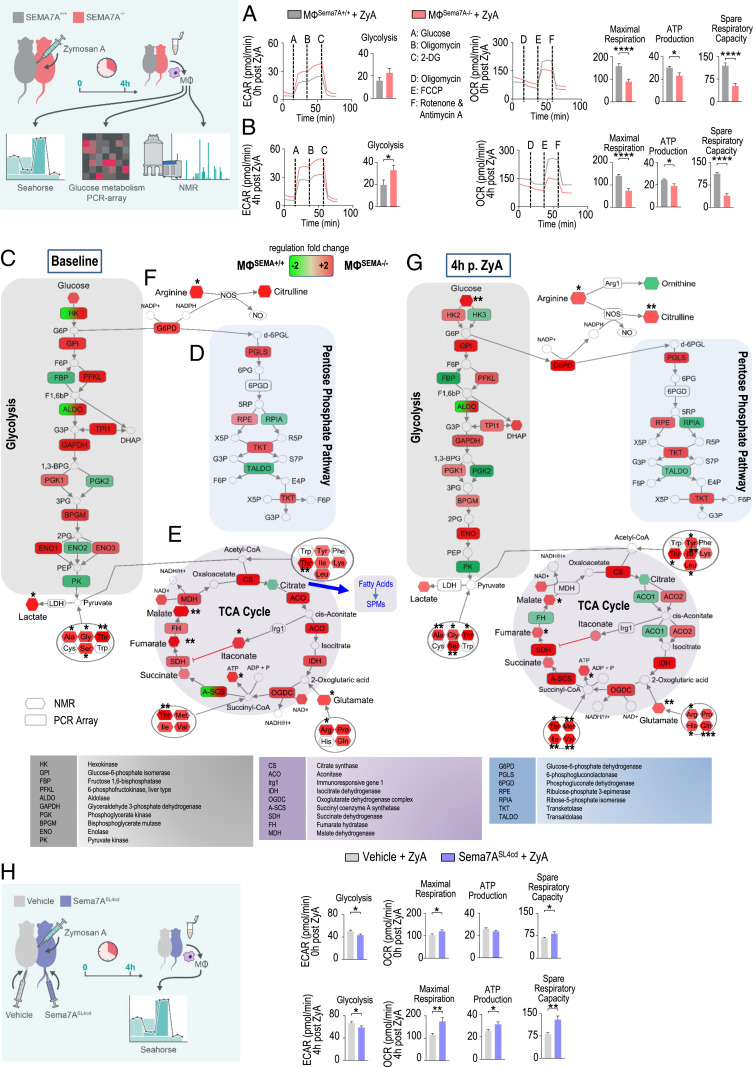
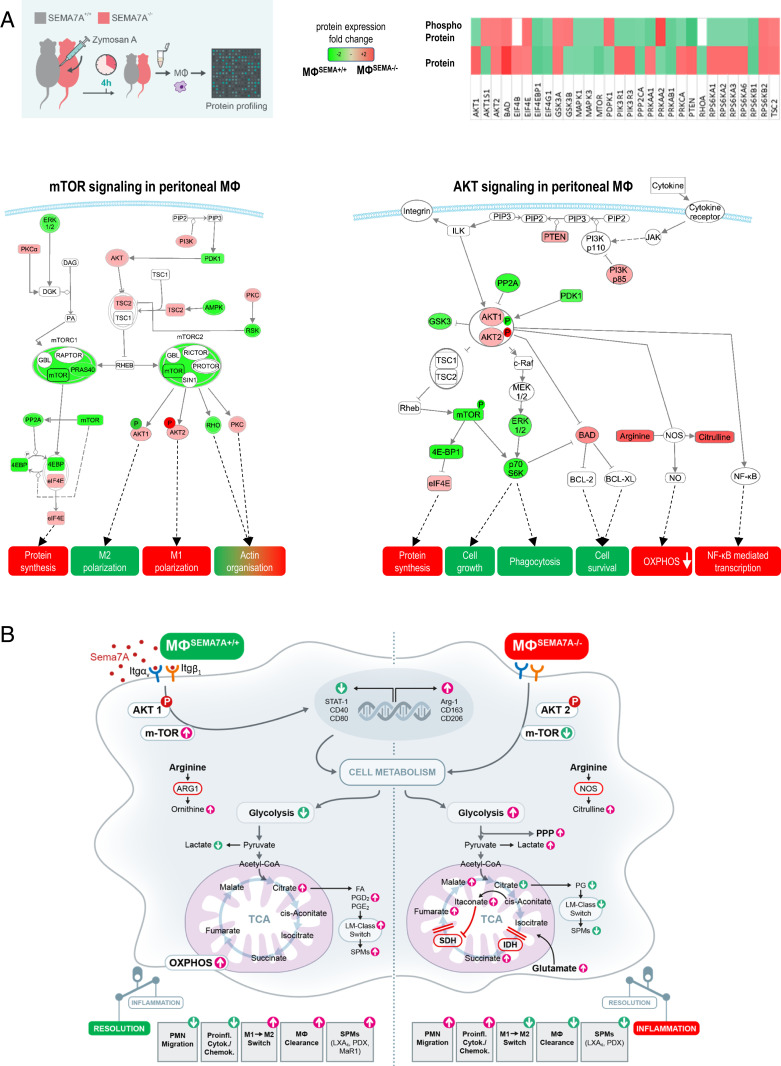
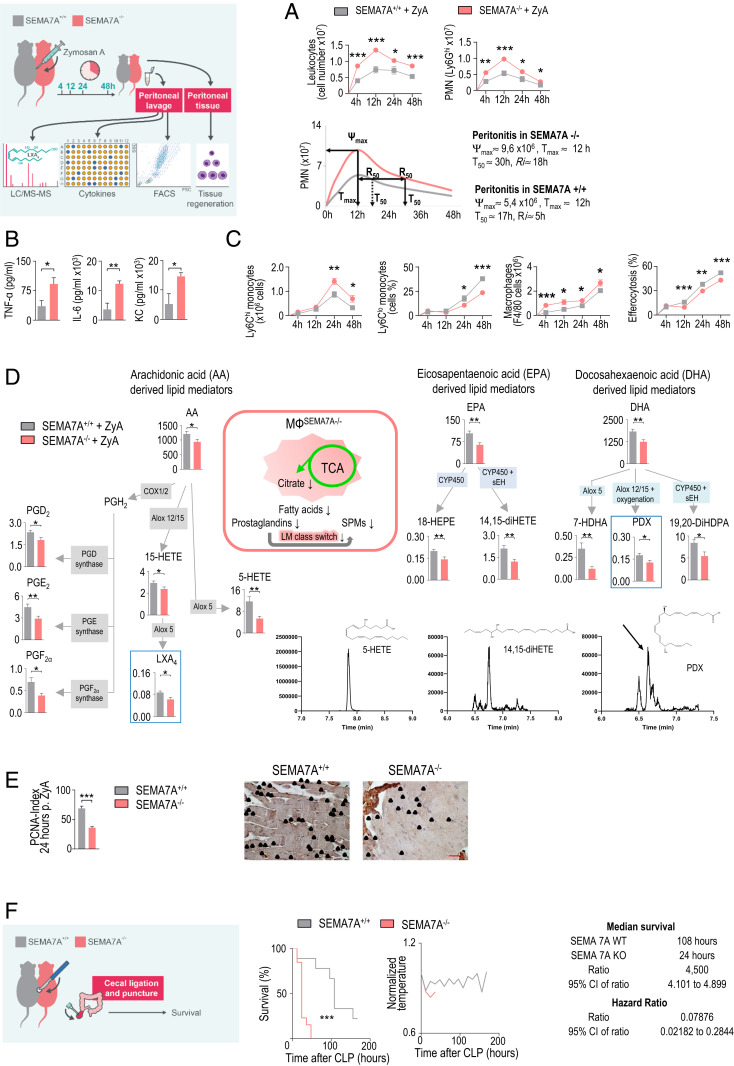
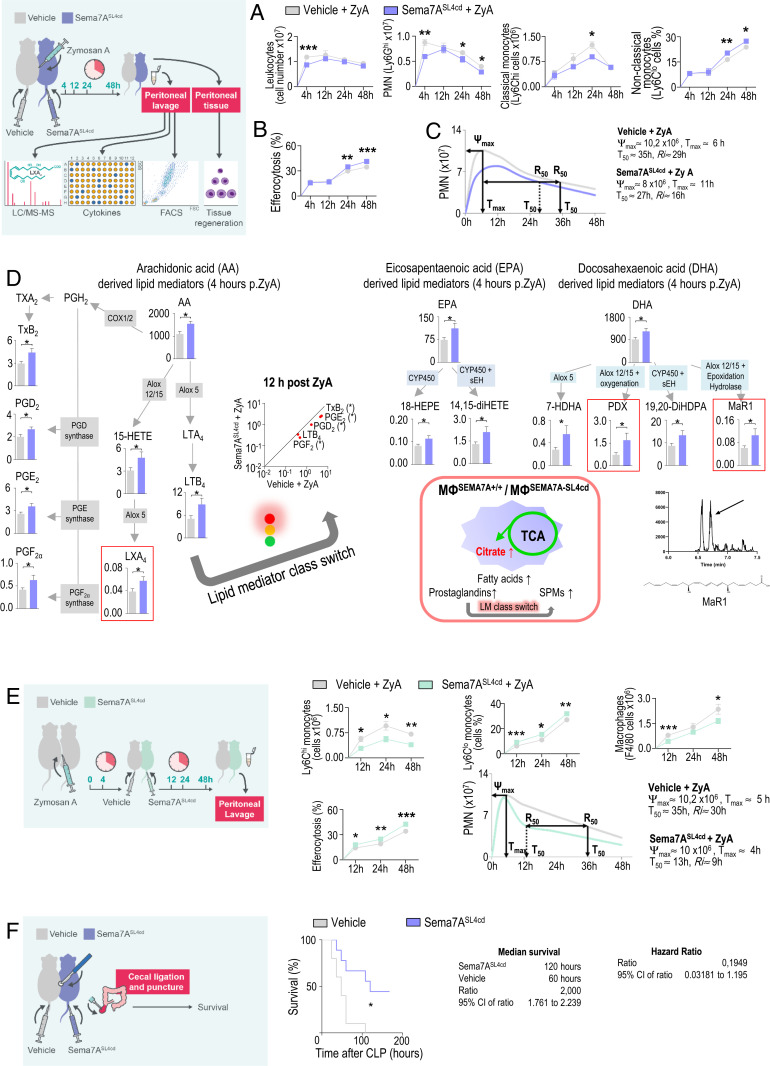
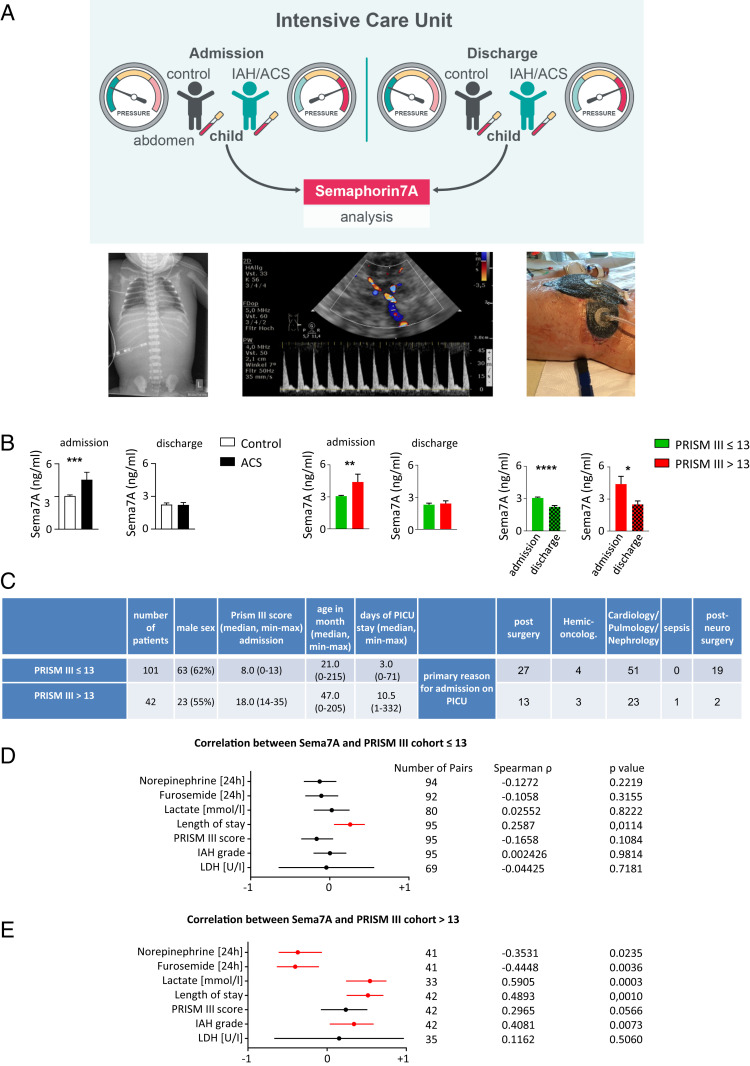
Endogenous mediators regulating acute inflammatory responses in both the induction and resolution phases of inflammatory processes are pivotal in host defense and tissue homeostasis. Recent studies have identified neuronal guidance proteins characterized in axonal development that display immunomodulatory functions. Here, we identify the neuroimmune guidance cue Semaphorin 7A (Sema7A), which appears to link macrophage (MΦ) metabolic remodeling to inflammation resolution. Sema7A orchestrated MΦ chemotaxis and chemokinesis, activated MΦ differentiation and polarization toward the proresolving M2 phenotype, and promoted leukocyte clearance. Peritoneal MΦSema7A-/- displayed metabolic reprogramming, characterized by reductions in fatty acid oxidation and oxidative phosphorylation, increases in glycolysis and the pentose phosphate pathway, and truncation of the tricarboxylic acid cycle, which resulted in increased levels of the intermediates succinate and fumarate. The low accumulation of citrate in MΦSema7A-/- correlated with the decreased synthesis of prostaglandins, leading to a reduced impact on lipid-mediator class switching and the generation of specialized pro resolving lipid mediators. Signaling network analysis indicated that Sema7A induced the metabolic reprogramming of MΦ by activating the mTOR- and AKT2-signaling pathways. Administration of Sema7ASL4cd orchestrated the resolution response to tissue homeostasis by shortening the resolution interval, promoting tissue protection in murine peritonitis, and enhancing survival in polymicrobial sepsis.
Keywords: Semaphorin 7A; inflammation; lipid mediator; metabolism; resolution.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Mazumdar C., Driggers E. M., Turka L. A., The untapped opportunity and challenge of immunometabolism: A new paradigm for drug discovery. Cell Metab. 31, 26–34 (2020). - PubMed
-
- Mirakaj V., Rosenberger P., Immunomodulatory functions of neuronal guidance proteins. Trends Immunol. 38, 444–456 (2017). - PubMed
-
- Suzuki K., Kumanogoh A., Kikutani H., Semaphorins and their receptors in immune cell interactions. Nat. Immunol. 9, 17–23 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
