

Cyclase-associated protein 2 (CAP2) controls MRTF-A localization and SRF activity in mouse embryonic fibroblasts
- PMID: 33637797
- PMCID: PMC7910472
- DOI: 10.1038/s41598-021-84213-w
Cyclase-associated protein 2 (CAP2) controls MRTF-A localization and SRF activity in mouse embryonic fibroblasts
Abstract
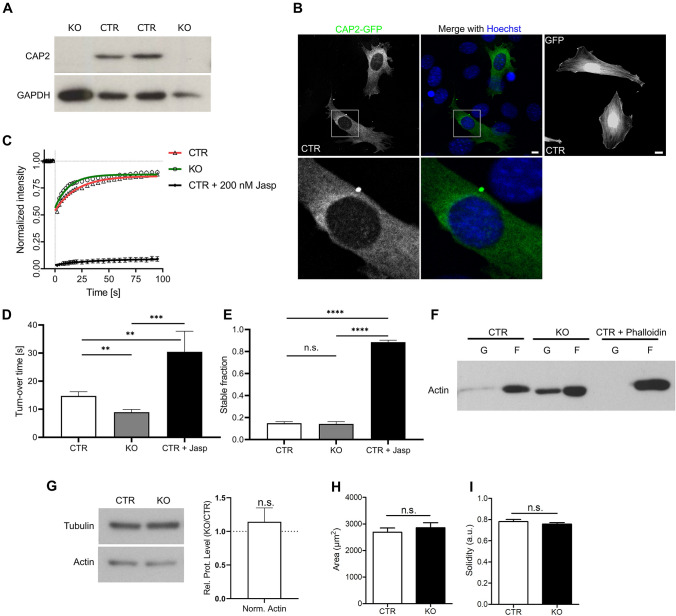
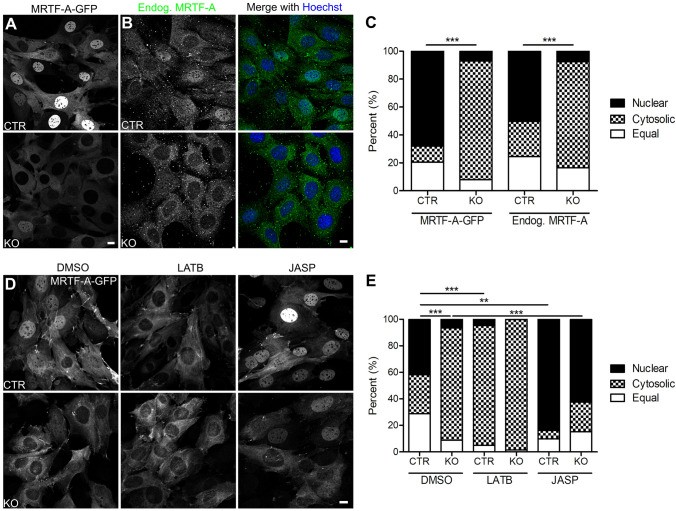
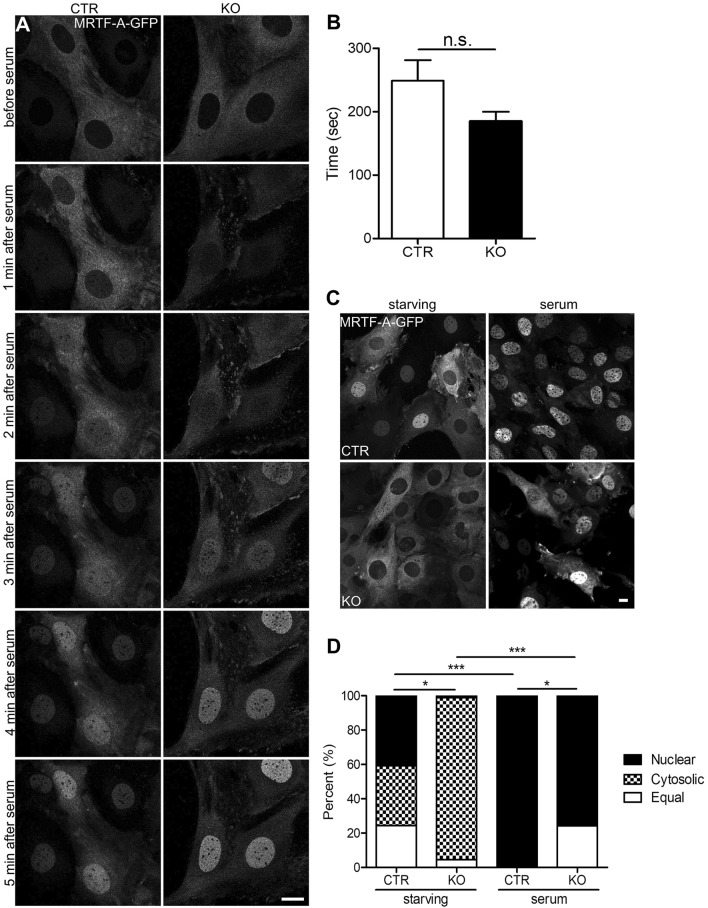
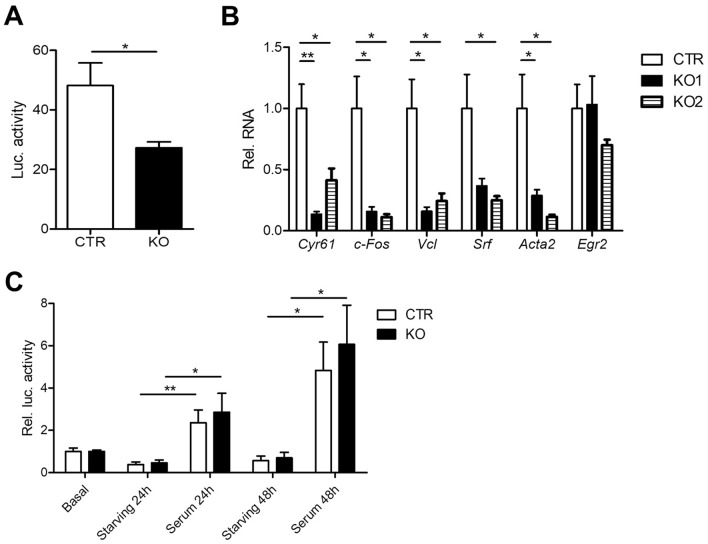
Recent studies identified cyclase-associated proteins (CAPs) as important regulators of actin dynamics that control assembly and disassembly of actin filaments (F-actin). While these studies significantly advanced our knowledge of their molecular functions, the physiological relevance of CAPs largely remained elusive. Gene targeting in mice implicated CAP2 in heart physiology and skeletal muscle development. Heart defects in CAP2 mutant mice were associated with altered activity of serum response factor (SRF), a transcription factor involved in multiple biological processes including heart function, but also skeletal muscle development. By exploiting mouse embryonic fibroblasts (MEFs) from CAP2 mutant mice, we aimed at deciphering the CAP2-dependent mechanism relevant for SRF activity. Reporter assays and mRNA quantification by qPCR revealed reduced SRF-dependent gene expression in mutant MEFs. Reduced SRF activity in CAP2 mutant MEFs was associated with altered actin turnover, a shift in the actin equilibrium towards monomeric actin (G-actin) as well as and reduced nuclear levels of myocardin-related transcription factor A (MRTF-A), a transcriptional SRF coactivator that is shuttled out of the nucleus and, hence, inhibited upon G-actin binding. Moreover, pharmacological actin manipulation with jasplakinolide restored MRTF-A distribution in mutant MEFs. Our data are in line with a model in which CAP2 controls the MRTF-SRF pathway in an actin-dependent manner. While MRTF-A localization and SRF activity was impaired under basal conditions, serum stimulation induced nuclear MRTF-A translocation and SRF activity in mutant MEFs similar to controls. In summary, our data revealed that in MEFs CAP2 controls basal MRTF-A localization and SRF activity, while it was dispensable for serum-induced nuclear MRTF-A translocation and SRF stimulation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
