

Macrophage migration inhibitory factor facilitates the therapeutic efficacy of mesenchymal stem cells derived exosomes in acute myocardial infarction through upregulating miR-133a-3p
- PMID: 33639970
- PMCID: PMC7916292
- DOI: 10.1186/s12951-021-00808-5
Macrophage migration inhibitory factor facilitates the therapeutic efficacy of mesenchymal stem cells derived exosomes in acute myocardial infarction through upregulating miR-133a-3p
Abstract
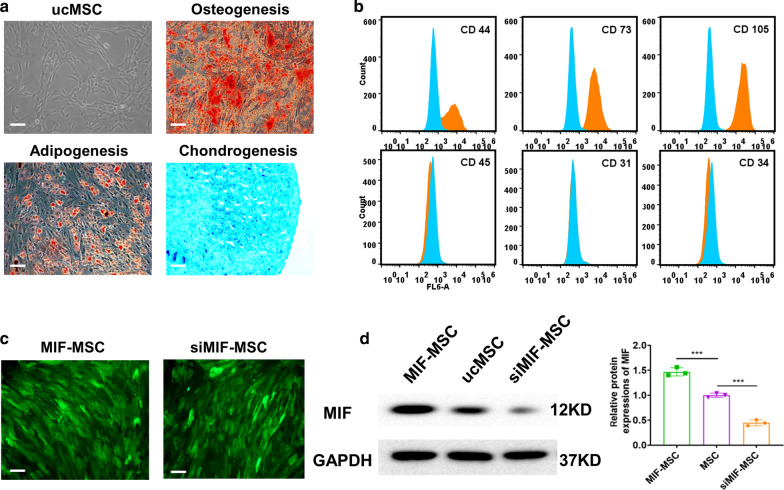
Background: Exosome transplantation is a promising cell-free therapeutic approach for the treatment of ischemic heart disease. The purpose of this study was to explore whether exosomes derived from Macrophage migration inhibitory factor (MIF) engineered umbilical cord MSCs (ucMSCs) exhibit superior cardioprotective effects in a rat model of AMI and reveal the mechanisms underlying it.
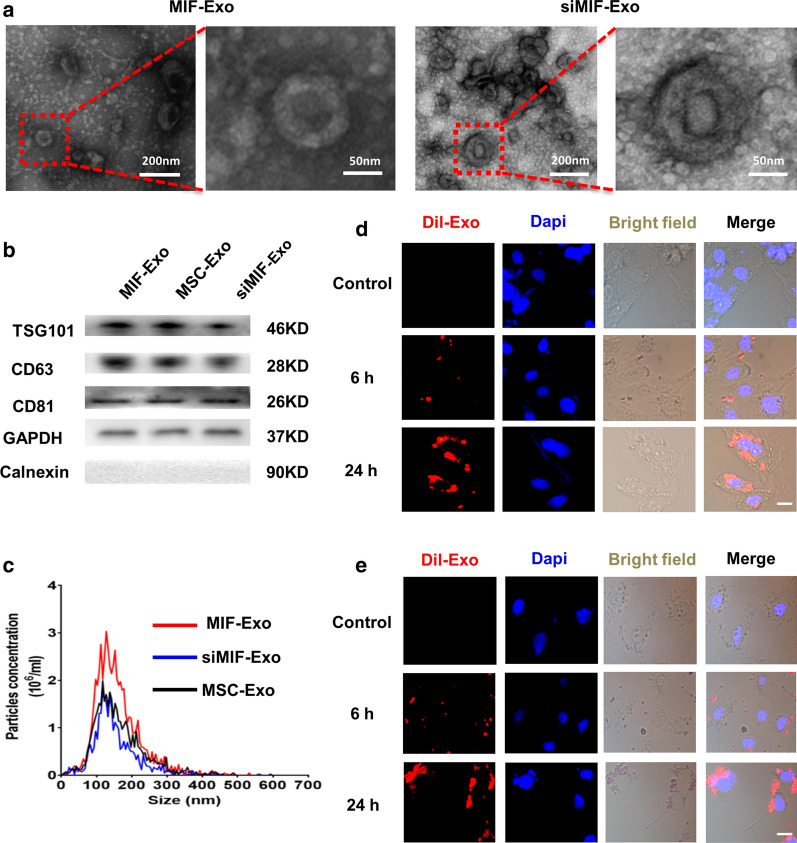
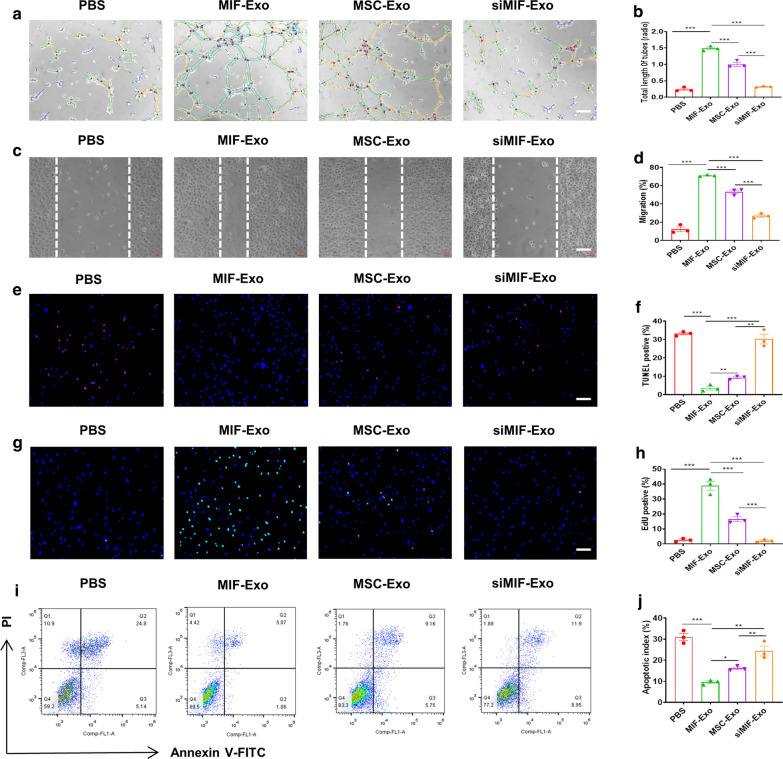
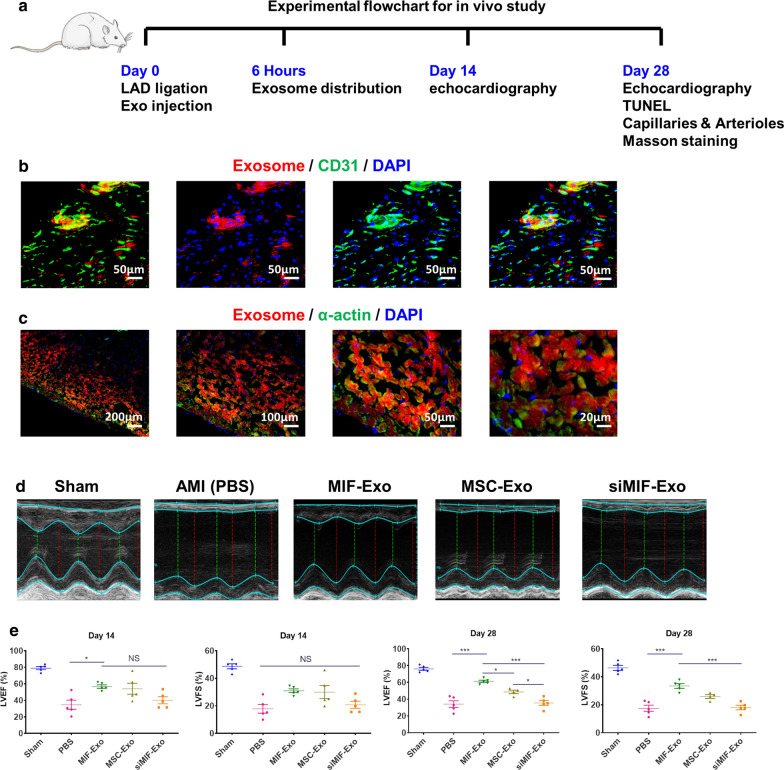
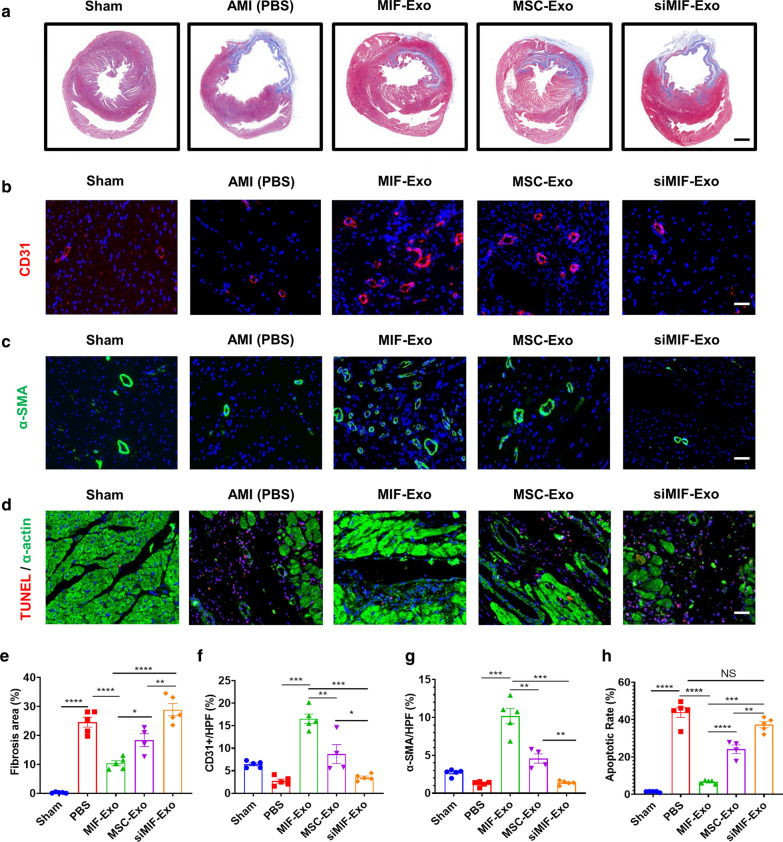
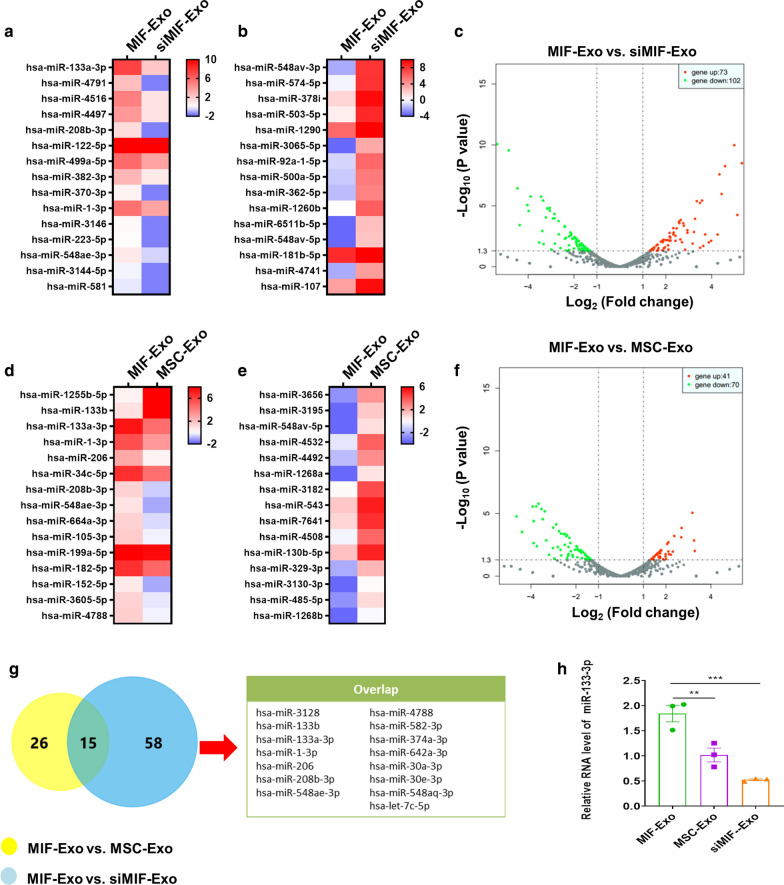
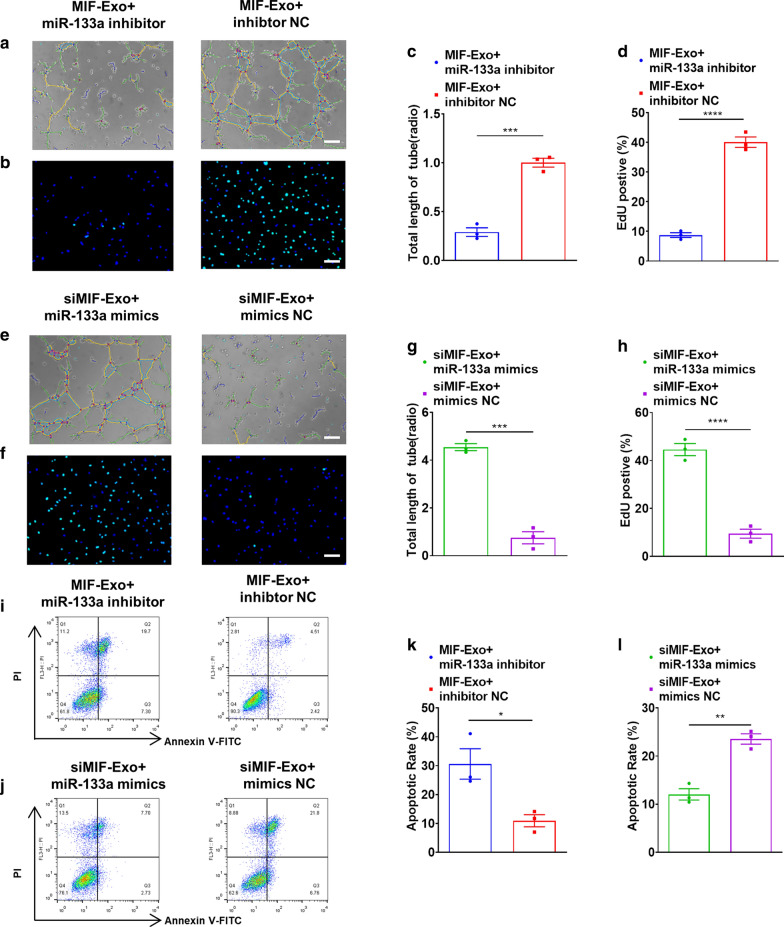
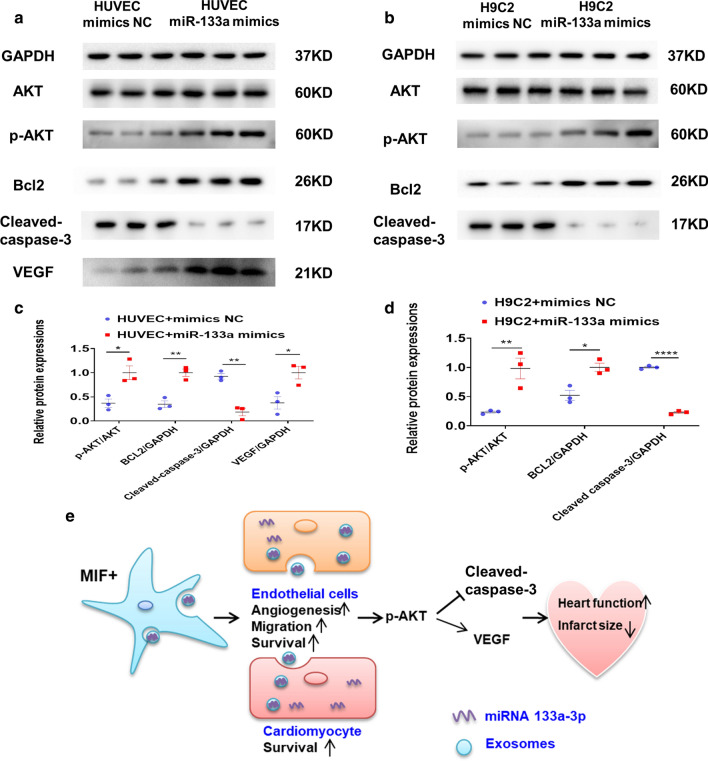
Results: Exosomes isolated from ucMSCs (MSC-Exo), MIF engineered ucMSCs (MIF-Exo) and MIF downregulated ucMSCs (siMIF-Exo) were used to investigate cellular protective function in human umbilical vein endothelial cells (HUVECs) and H9C2 cardiomyocytes under hypoxia and serum deprivation (H/SD) and infarcted hearts in rats. Compared with MSC-Exo and siMIF-Exo, MIF-Exo significantly enhanced proliferation, migration, and angiogenesis of HUVECs and inhibited H9C2 cardiomyocyte apoptosis under H/SD in vitro. MIF-Exo also significantly inhibited cardiomyocyte apoptosis, reduced fibrotic area, and improved cardiac function as measured by echocardiography in infarcted rats in vivo. Exosomal miRNAs sequencing and qRT-PCR confirmed miRNA-133a-3p significantly increased in MIF-Exo. The biological effects of HUVECs and H9C2 cardiomyocytes were attenuated with incubation of MIF-Exo and miR-133a-3p inhibitors. These effects were accentuated with incubation of siMIF-Exo and miR-133a-3p mimics that increased the phosphorylation of AKT protein in these cells.
Conclusion: MIF-Exo can provide cardioprotective effects by promoting angiogenesis, inhibiting apoptosis, reducing fibrosis, and preserving heart function in vitro and in vivo. The mechanism in the biological activities of MIF-Exo involves miR-133a-3p and the downstream AKT signaling pathway.
Keywords: Exosomes; Macrophage migration inhibitory factor; MiR-133a-3p; Myocardial infarction; UcMSCs.
Conflict of interest statement
The authors have declared that no competing interest exists.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
