

Importin-αs are required for the nuclear localization and function of the Plasmopara viticola effector PvAVH53
- PMID: 33642571
- PMCID: PMC7917100
- DOI: 10.1038/s41438-021-00482-6
Importin-αs are required for the nuclear localization and function of the Plasmopara viticola effector PvAVH53
Abstract
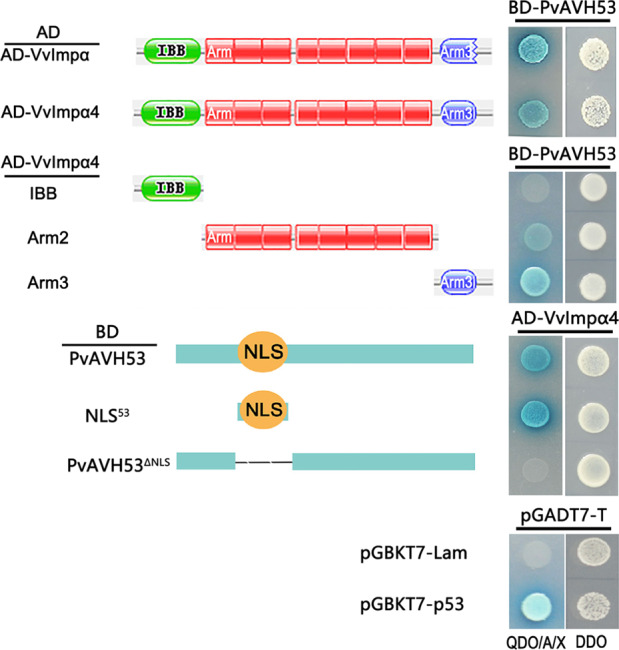
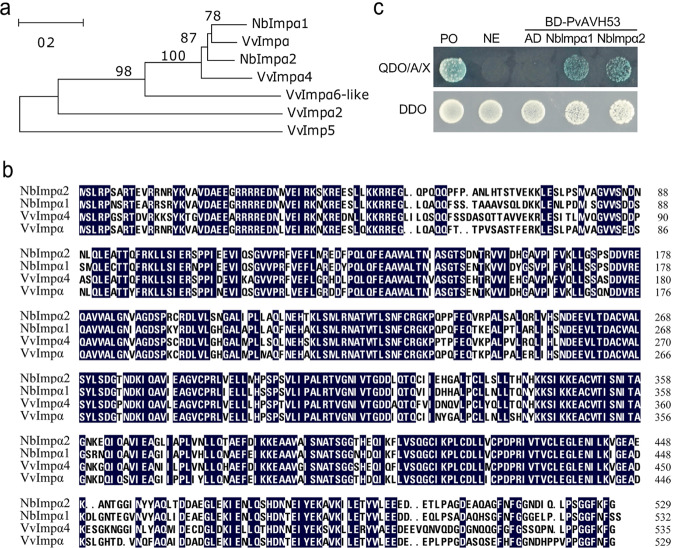
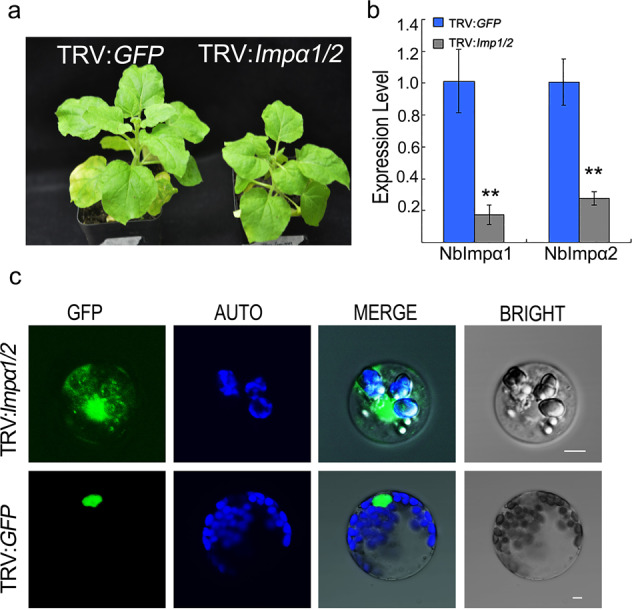
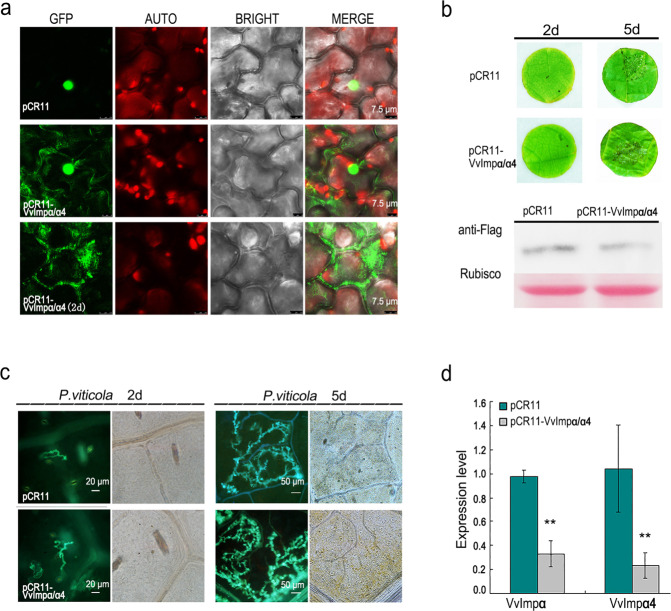
Plant pathogenic oomycetes deliver a troop of effector proteins into the nucleus of host cells to manipulate plant cellular immunity and promote colonization. Recently, researchers have focused on identifying how effectors are transferred into the host cell nucleus, as well as the identity of the nuclear targets. In this study, we found that the RxLR effector PvAVH53 from the grapevine (Vitis vinifera) oomycete pathogen Plasmopara viticola physically interacts with grapevine nuclear import factor importin alphas (VvImpα and VvImpα4), localizes to the nucleus and triggers cell death when transiently expressed in tobacco (Nicotiana benthamiana) cells. Deletion of a nuclear localization signal (NLS) sequence from PvAVH53 or addition of a nuclear export signal (NES) sequence disrupted the nuclear localization of PvAVH53 and attenuated its ability to trigger cell death. Suppression of two tobacco importin-α genes, namely, NbImp-α1 and NbImp-α2, by virus-induced gene silencing (VIGS) also disrupted the nuclear localization and ability of PvAVH53 to induce cell death. Likewise, we transiently silenced the expression of VvImpα/α4 in grape through CRISPR/Cas13a, which has been reported to target RNA in vivo. Finally, we found that attenuating the expression of the Importin-αs genes resulted in increased susceptibility to the oomycete pathogen Phytophthora capsici in N. benthamiana and P. viticola in V. vinifera. Our results demonstrate that importin-αs are required for the nuclear localization and function of PvAVH53 and are essential for host innate immunity. The findings provide insight into the functions of importin-αs in grapevine against downy mildew.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
Grants and funding
- Grant No. 31471844/National Natural Science Foundation of China (National Science Foundation of China)
- Grants No. 31471844/National Natural Science Foundation of China (National Science Foundation of China)
- Grants No. 31471844/National Natural Science Foundation of China (National Science Foundation of China)
- Grants No. 31471844/National Natural Science Foundation of China (National Science Foundation of China)
- Grant No. 201203075-08/Special Fund for Agro-scientific Research in the Public Interest (Agro-Scientific Research in the Public Interest of China)
LinkOut - more resources
Full Text Sources
Other Literature Sources
