

Place vs. Response Learning: History, Controversy, and Neurobiology
- PMID: 33643005
- PMCID: PMC7904695
- DOI: 10.3389/fnbeh.2020.598570
Place vs. Response Learning: History, Controversy, and Neurobiology
Abstract
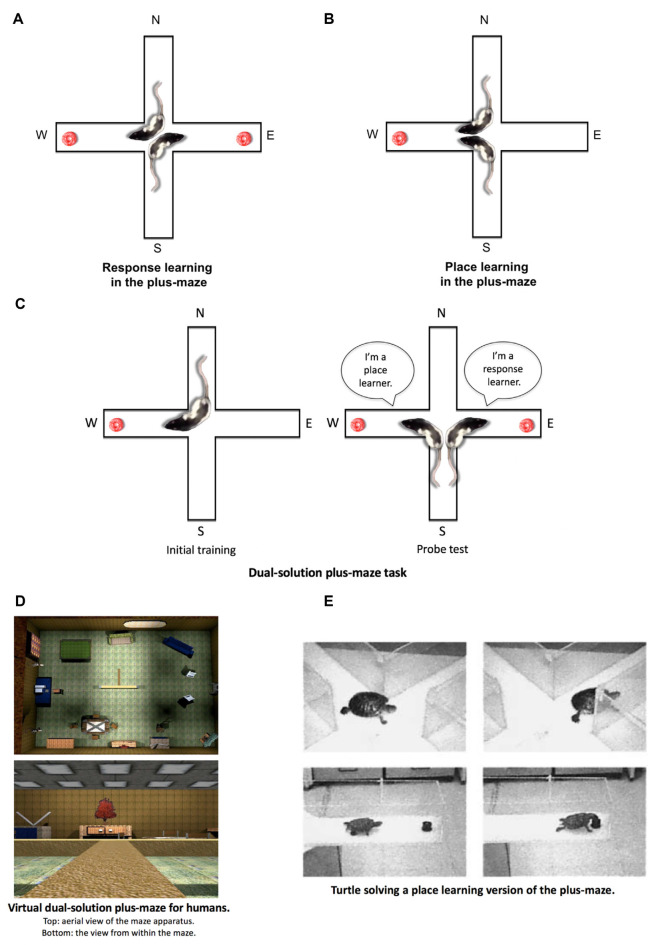
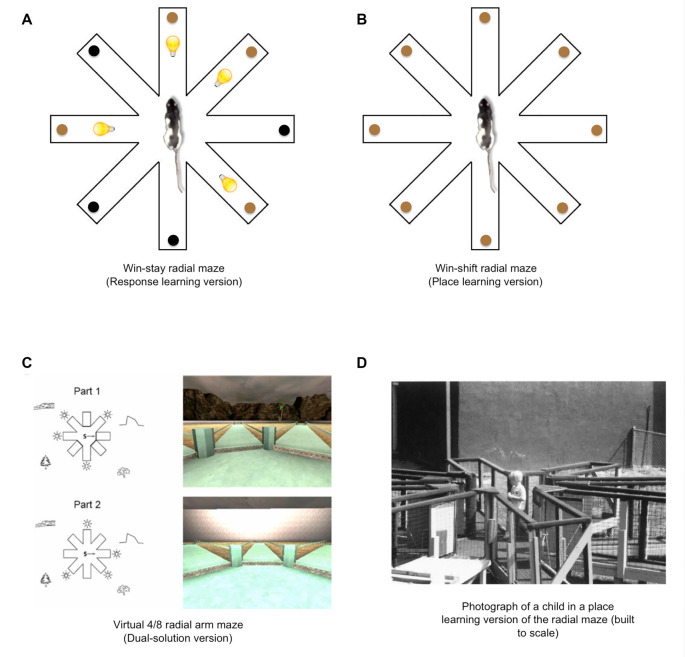
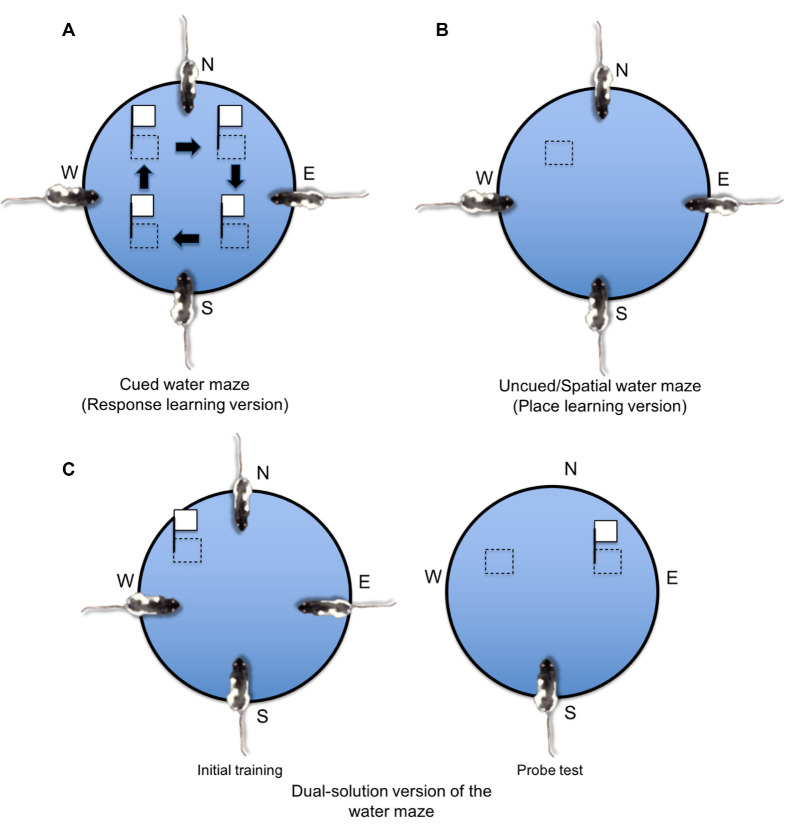
The present article provides a historical review of the place and response learning plus-maze tasks with a focus on the behavioral and neurobiological findings. The article begins by reviewing the conflict between Edward C. Tolman's cognitive view and Clark L. Hull's stimulus-response (S-R) view of learning and how the place and response learning plus-maze tasks were designed to resolve this debate. Cognitive learning theorists predicted that place learning would be acquired faster than response learning, indicating the dominance of cognitive learning, whereas S-R learning theorists predicted that response learning would be acquired faster, indicating the dominance of S-R learning. Here, the evidence is reviewed demonstrating that either place or response learning may be dominant in a given learning situation and that the relative dominance of place and response learning depends on various parametric factors (i.e., amount of training, visual aspects of the learning environment, emotional arousal, et cetera). Next, the neurobiology underlying place and response learning is reviewed, providing strong evidence for the existence of multiple memory systems in the mammalian brain. Research has indicated that place learning is principally mediated by the hippocampus, whereas response learning is mediated by the dorsolateral striatum. Other brain regions implicated in place and response learning are also discussed in this section, including the dorsomedial striatum, amygdala, and medial prefrontal cortex. An exhaustive review of the neurotransmitter systems underlying place and response learning is subsequently provided, indicating important roles for glutamate, dopamine, acetylcholine, cannabinoids, and estrogen. Closing remarks are made emphasizing the historical importance of the place and response learning tasks in resolving problems in learning theory, as well as for examining the behavioral and neurobiological mechanisms of multiple memory systems. How the place and response learning tasks may be employed in the future for examining extinction, neural circuits of memory, and human psychopathology is also briefly considered.
Keywords: dorsal striatum; habit; hippocampus; memory systems; place learning; response learning; spatial memory; stimulus-response learning.
Copyright © 2021 Goodman.
Conflict of interest statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


Similar articles
-
The amygdala and emotional modulation of competition between cognitive and habit memory.Behav Brain Res. 2008 Nov 3;193(1):126-31. doi: 10.1016/j.bbr.2008.05.002. Epub 2008 May 8. Behav Brain Res. 2008. PMID: 18565602
-
Exhumed from thought: basal ganglia and response learning in the plus-maze.Behav Brain Res. 2009 Apr 12;199(1):24-31. doi: 10.1016/j.bbr.2008.12.013. Epub 2008 Dec 14. Behav Brain Res. 2009. PMID: 19133296 Review.
-
The role of the dorsal striatum in extinction: A memory systems perspective.Neurobiol Learn Mem. 2018 Apr;150:48-55. doi: 10.1016/j.nlm.2018.02.028. Epub 2018 Mar 1. Neurobiol Learn Mem. 2018. PMID: 29501803 Review.
-
The influence of cannabinoids on learning and memory processes of the dorsal striatum.Neurobiol Learn Mem. 2015 Nov;125:1-14. doi: 10.1016/j.nlm.2015.06.008. Epub 2015 Jun 17. Neurobiol Learn Mem. 2015. PMID: 26092091 Review.
-
The dorsolateral striatum selectively mediates extinction of habit memory.Neurobiol Learn Mem. 2016 Dec;136:54-62. doi: 10.1016/j.nlm.2016.09.012. Epub 2016 Sep 20. Neurobiol Learn Mem. 2016. PMID: 27663194
Cited by
-
Navigation task and action space drive the emergence of egocentric and allocentric spatial representations.PLoS Comput Biol. 2022 Oct 31;18(10):e1010320. doi: 10.1371/journal.pcbi.1010320. eCollection 2022 Oct. PLoS Comput Biol. 2022. PMID: 36315587 Free PMC article.
-
Chronic corticosterone administration in adolescence enhances dorsolateral striatum-dependent learning in adulthood.Front Behav Neurosci. 2022 Aug 12;16:970304. doi: 10.3389/fnbeh.2022.970304. eCollection 2022. Front Behav Neurosci. 2022. PMID: 36035016 Free PMC article.
-
Reply to Calvo, Raja, and Segundo-Ortin, "Don't jump the gun quite yet: aiming for the true target in plant neurobiology research".Protoplasma. 2025 Mar;262(2):277-278. doi: 10.1007/s00709-024-02007-z. Epub 2024 Nov 15. Protoplasma. 2025. PMID: 39545947
-
Function and biochemistry of the dorsolateral prefrontal cortex during placebo analgesia: how the certainty of prior experiences shapes endogenous pain relief.Cereb Cortex. 2023 Aug 23;33(17):9822-9834. doi: 10.1093/cercor/bhad247. Cereb Cortex. 2023. PMID: 37415068 Free PMC article.
-
Chronic alcohol consumption shifts learning strategies and synaptic plasticity from hippocampus to striatum-dependent pathways.Front Psychiatry. 2023 May 26;14:1129030. doi: 10.3389/fpsyt.2023.1129030. eCollection 2023. Front Psychiatry. 2023. PMID: 37304443 Free PMC article.
References
-
- Adams C. D. (1982). Variations in the sensitivity of instrumental responding to reinforcer devaluation. Q. J. Exp. Psychol. 34, 77–98. 10.1080/14640748208400878 - DOI
-
- Adams C. D., Dickinson A. (1981a). Instrumental responding following reinforcer devaluation. Q. J. Exp. Psychol. 33, 109–121. 10.1080/14640748108400816 - DOI
-
- Adams C. D., Dickinson A. (1981b). “Actions and habits: variations in associative representations during instrumental learning,” in Information Processing in Animals: Memory Mechanisms, eds Spear N. E., Miller R. R. (Hillsdale, NJ: Erlbaum; ), 143–165.
-
- Almey A., Cannell E., Bertram K., Filardo E., Milner T. A., Brake W. G. (2014). Medial prefrontal cortical estradiol rapidly alters memory system bias in female rats: ultrastructural analysis reveals membrane-associated estrogen receptors as potential mediators. Endocrinology 155, 4422–4432. 10.1210/en.2014-1463 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials