

Suppression of Calcineurin Enhances the Toxicity of Cry1Ac to Helicoverpa armigera
- PMID: 33643268
- PMCID: PMC7904703
- DOI: 10.3389/fmicb.2021.634619
Suppression of Calcineurin Enhances the Toxicity of Cry1Ac to Helicoverpa armigera
Abstract
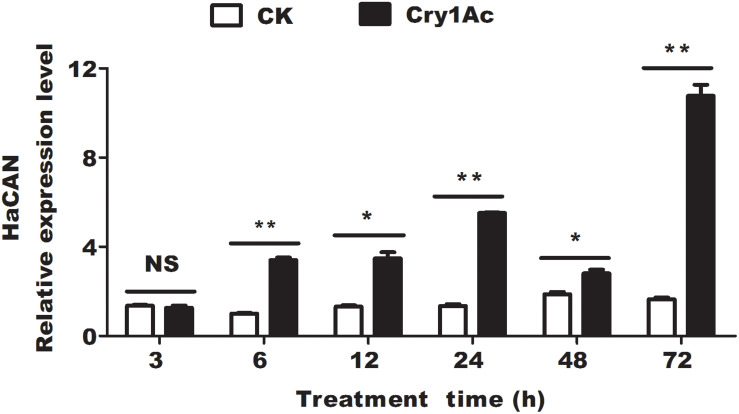
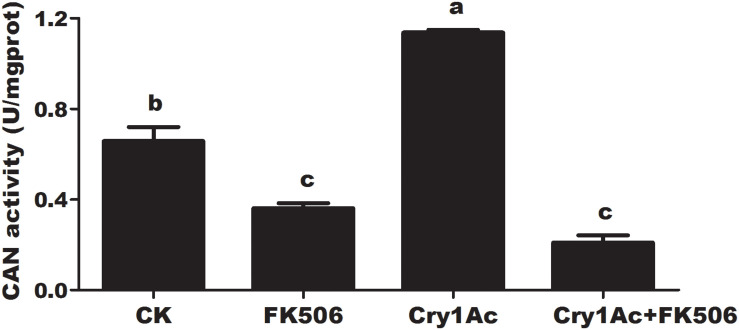
Insect resistance to Bacillus thuringiensis (Bt) insecticidal proteins has rapidly evolved with the expansion of the planting area of transgenic Bt crops. Pyramiding RNA interference (RNAi) and Bt in crops is urgently needed to counter the rapid increase in pest resistance. The ideal "pyramid" strategy simultaneously targets different action pathways that exert synergetic effects on each other. Here, we identified a dephosphatase, namely, Helicoverpa armigera calcineurin (HaCAN), which might enhance the insecticidal activity of Cry1Ac against Helicoverpa armigera by regulating immune gene expression via dephosphatase activity, but not by acting as a receptor. Notably, blocking enzyme activity or knocking down endogenous HaCAN significantly promoted the enhancement in Cry1Ac toxicity to insect larvae and cells. Correspondingly, the increase in HaCAN activity reduced the cytotoxicity of Cry1Ac as shown by the heterologous expression of HaCAN. Our results provide a probable that HaCAN is an important candidate gene for pyramiding RNAi and Cry1Ac crops to control cotton bollworm.
Keywords: Cry1Ac; Helicoverpa armigera; calcineurin; cell toxicity; membrane yeast two-hybrid.
Copyright © 2021 Wei, Yao, Yang, Liu, Zhou, Cen, Liu, Du, Tang and An.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Ballesteros-Martinez C., Mendez-Barbero N., Montalvo-Yuste A., Jensen B. M., Gomez-Cardenosa A., Klitfod L. (2017). Endothelial regulator of calcineurin 1 promotes barrier integrity and modulates histamine-induced barrier dysfunction in anaphylaxis. Front. Immunol. 8:1323. 10.3389/fimmu.2017.01323 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources