

P. aeruginosa Mediated Necroptosis in Mouse Tumor Cells Induces Long-Lasting Systemic Antitumor Immunity
- PMID: 33643911
- PMCID: PMC7908819
- DOI: 10.3389/fonc.2020.610651
P. aeruginosa Mediated Necroptosis in Mouse Tumor Cells Induces Long-Lasting Systemic Antitumor Immunity
Abstract
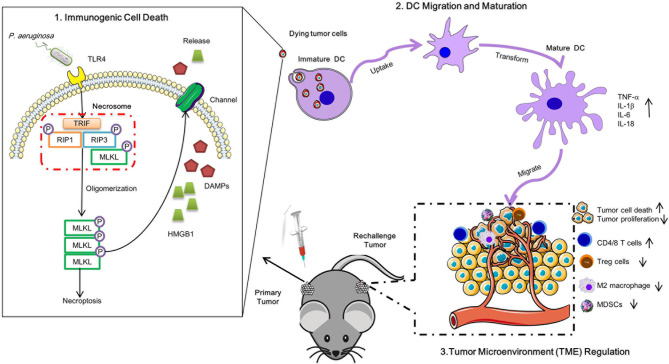
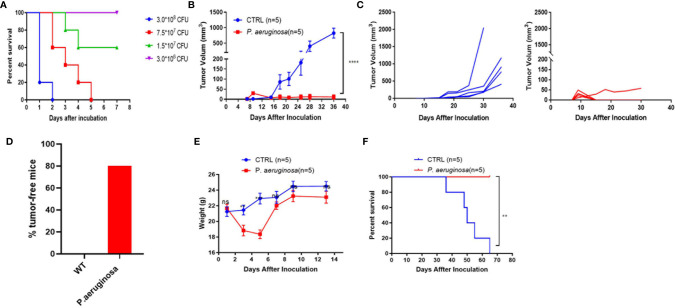
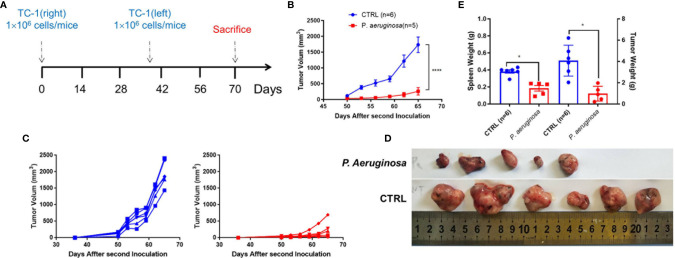
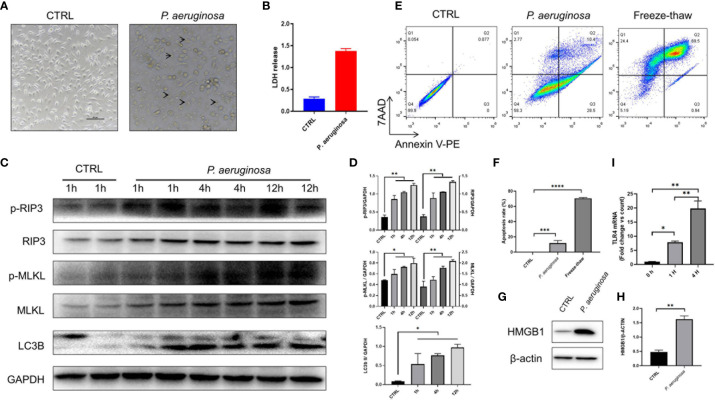
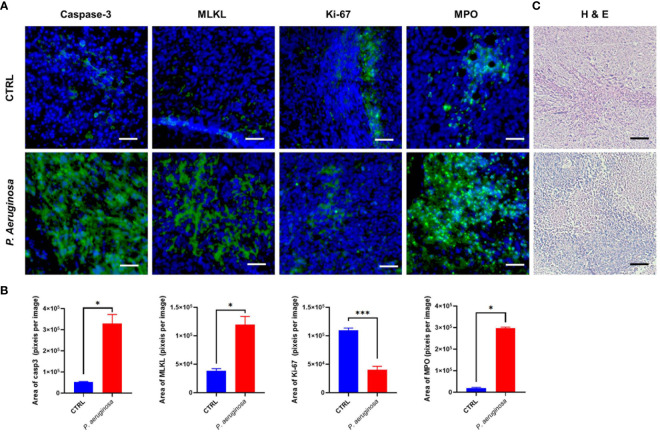
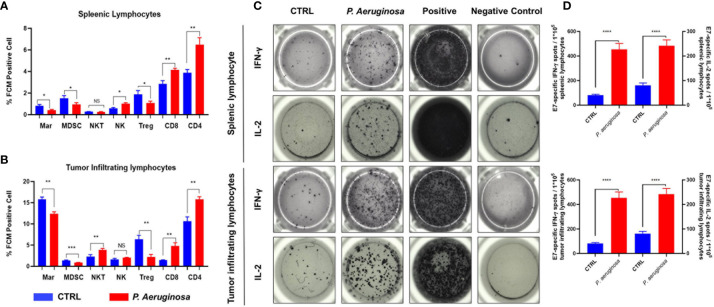
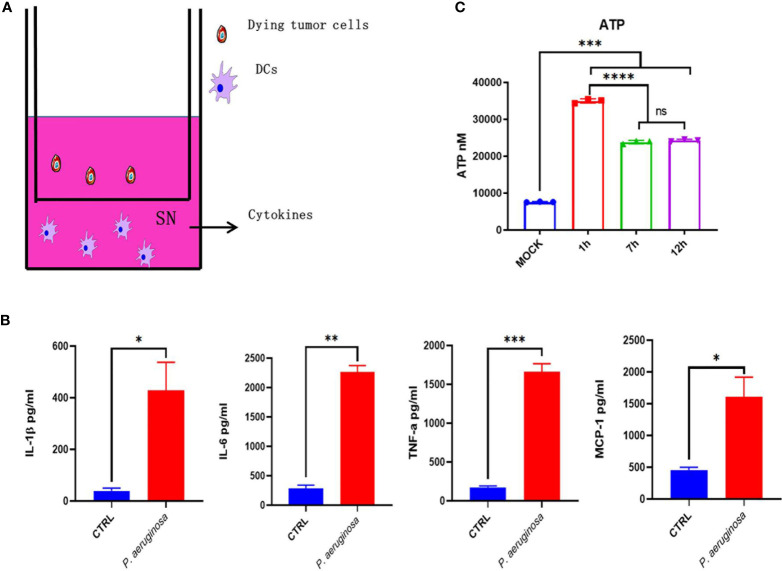
Necroptosis is a form of programmed cell death (PCD) characterized by RIP3 mediated MLKL activation and increased membrane permeability via MLKL oligomerization. Tumor cell immunogenic cell death (ICD) has been considered to be essential for the anti-tumor response, which is associated with DC recruitment, activation, and maturation. In this study, we found that P. aeruginosa showed its potential to suppress tumor growth and enable long-lasting anti-tumor immunity in vivo. What's more, phosphorylation- RIP3 and MLKL activation induced by P. aeruginosa infection resulted in tumor cell necrotic cell death and HMGB1 production, indicating that P. aeruginosa can cause immunogenic cell death. The necrotic cell death can further drive a robust anti-tumor response via promoting tumor cell death, inhibiting tumor cell proliferation, and modulating systemic immune responses and local immune microenvironment in tumor. Moreover, dying tumor cells killed by P. aeruginosa can catalyze DC maturation, which enhanced the antigen-presenting ability of DC cells. These findings demonstrate that P. aeruginosa can induce immunogenic cell death and trigger a robust long-lasting anti-tumor response along with reshaping tumor microenvironment.
Keywords: P. aeruginosa; antitumor immunity; dying tumor cell; necroptosis; tumor microenvironment.
Copyright © 2021 Qi, He, Jin, Yang, Bai, Liu and Ma.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous