

Sealing holes in cellular membranes
- PMID: 33644904
- PMCID: PMC8013788
- DOI: 10.15252/embj.2020106922
Sealing holes in cellular membranes
Abstract
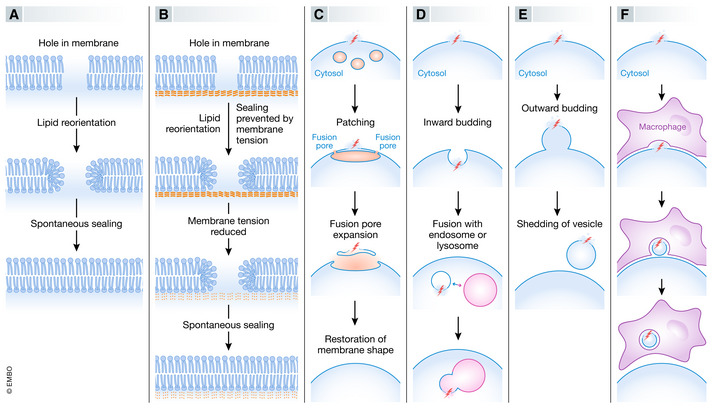
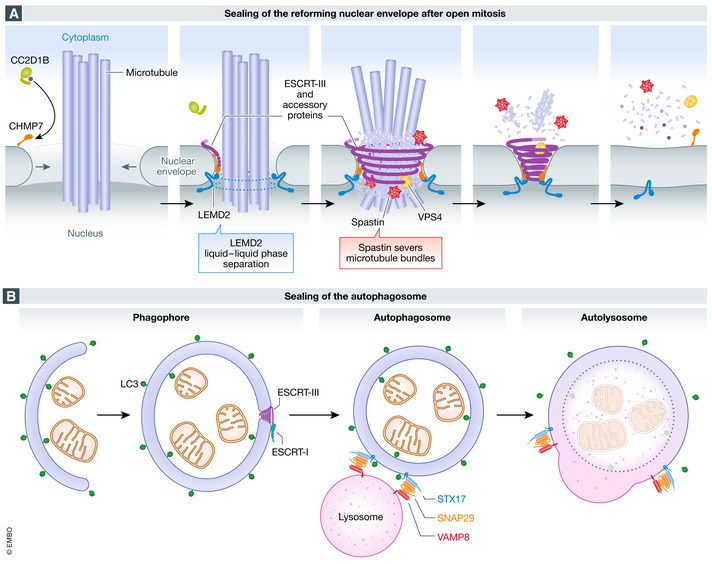
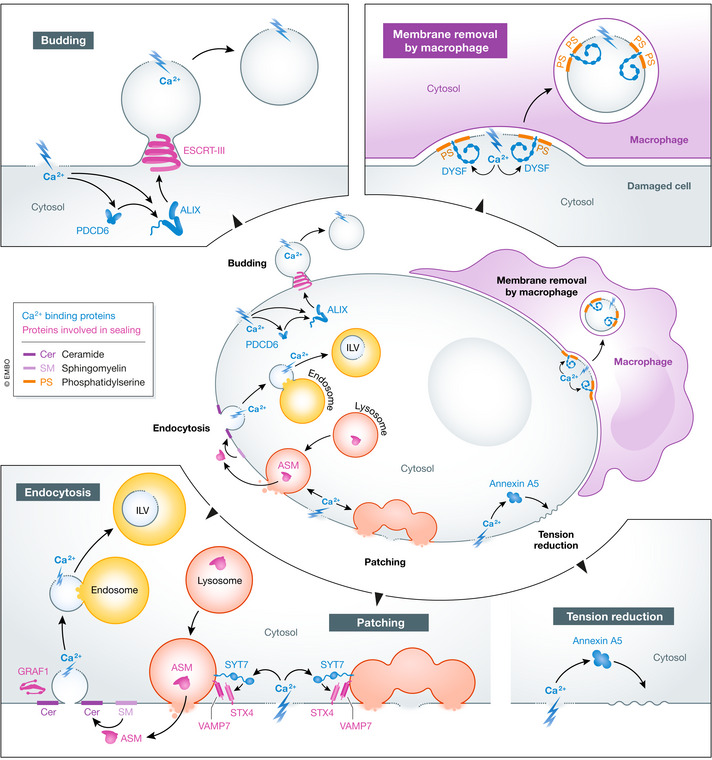
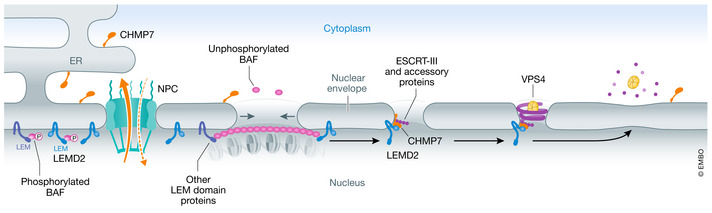
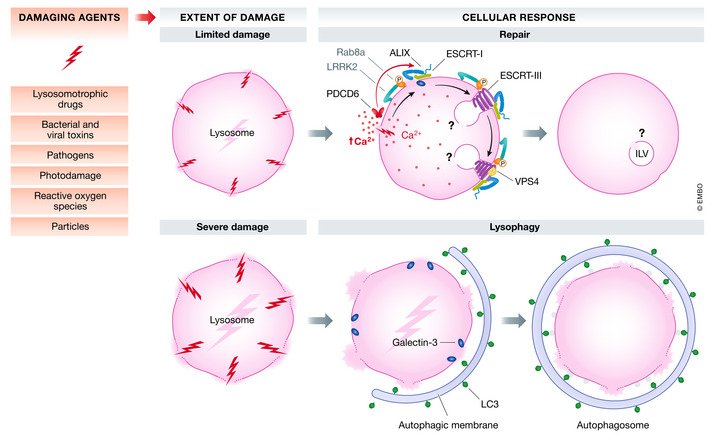
The compartmentalization of eukaryotic cells, which is essential for their viability and functions, is ensured by single or double bilayer membranes that separate the cell from the exterior and form boundaries between the cell's organelles and the cytosol. Nascent nuclear envelopes and autophagosomes, which both are enveloped by double membranes, need to be sealed during the late stage of their biogenesis. On the other hand, the integrity of cellular membranes such as the plasma membrane, lysosomes and the nuclear envelope can be compromised by pathogens, chemicals, radiation, inflammatory responses and mechanical stress. There are cellular programmes that restore membrane integrity after injury. Here, we review cellular mechanisms that have evolved to maintain membrane integrity during organelle biogenesis and after injury, including membrane scission mediated by the endosomal sorting complex required for transport (ESCRT), vesicle patching and endocytosis.
Keywords: ESCRT; autophagy; endocytosis; lysosome; membrane repair.
© 2021 The Authors. Published under the terms of the CC BY NC ND 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Berg TO, Fengsrud M, Stromhaug PE, Berg T, Seglen PO (1998) Isolation and characterization of rat liver amphisomes ‐ Evidence for fusion of autophagosomes with both early and late endosomes. J Biol Chem 273: 21883–21892 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
