

Supramolecular associations between atypical oxidative phosphorylation complexes of Euglena gracilis
- PMID: 33646522
- PMCID: PMC8124061
- DOI: 10.1007/s10863-021-09882-8
Supramolecular associations between atypical oxidative phosphorylation complexes of Euglena gracilis
Erratum in
-
Correction to: Supramolecular associations between atypical oxidative phosphorylation complexes of Euglena gracilis.J Bioenerg Biomembr. 2021 Jun;53(3):365. doi: 10.1007/s10863-021-09890-8. J Bioenerg Biomembr. 2021. PMID: 33978869 Free PMC article. No abstract available.
Abstract
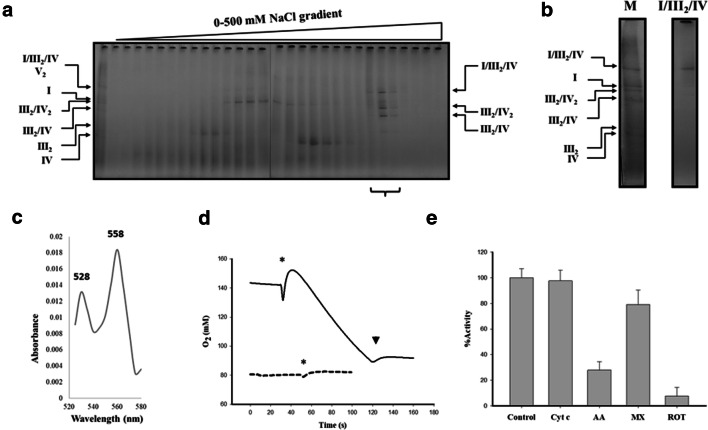
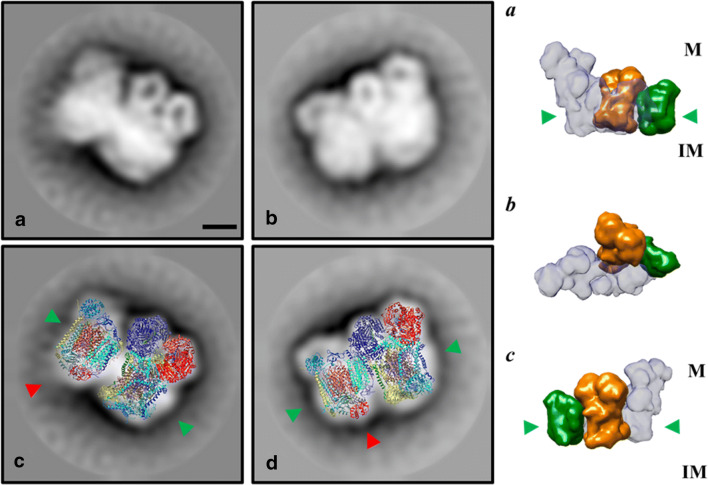
In vivo associations of respiratory complexes forming higher supramolecular structures are generally accepted nowadays. Supercomplexes (SC) built by complexes I, III and IV and the so-called respirasome (I/III2/IV) have been described in mitochondria from several model organisms (yeasts, mammals and green plants), but information is scarce in other lineages. Here we studied the supramolecular associations between the complexes I, III, IV and V from the secondary photosynthetic flagellate Euglena gracilis with an approach that involves the extraction with several mild detergents followed by native electrophoresis. Despite the presence of atypical subunit composition and additional structural domains described in Euglena complexes I, IV and V, canonical associations into III2/IV, III2/IV2 SCs and I/III2/IV respirasome were observed together with two oligomeric forms of the ATP synthase (V2 and V4). Among them, III2/IV SC could be observed by electron microscopy. The respirasome was further purified by two-step liquid chromatography and showed in-vitro oxygen consumption independent of the addition of external cytochrome c.
Keywords: Euglena gracilis; F1FO ATP synthase; Mitochondrial supercomplexes; Oligomeric complex V; Oxidative phosphorylation; Respirasome.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Oxidative phosphorylation supercomplexes and respirasome reconstitution of the colorless alga Polytomella sp.Biochim Biophys Acta Bioenerg. 2018 Jun;1859(6):434-444. doi: 10.1016/j.bbabio.2018.03.004. Epub 2018 Mar 11. Biochim Biophys Acta Bioenerg. 2018. PMID: 29540299
-
The atypical subunit composition of respiratory complexes I and IV is associated with original extra structural domains in Euglena gracilis.Sci Rep. 2018 Jun 26;8(1):9698. doi: 10.1038/s41598-018-28039-z. Sci Rep. 2018. PMID: 29946152 Free PMC article.
-
Respiratory chain supercomplexes in plant mitochondria.Plant Physiol Biochem. 2004 Dec;42(12):937-42. doi: 10.1016/j.plaphy.2004.09.010. Epub 2005 Jan 21. Plant Physiol Biochem. 2004. PMID: 15707832 Review.
-
Mitochondrial respirasome works as a single unit and the cross-talk between complexes I, III2 and IV stimulates NADH dehydrogenase activity.Biochim Biophys Acta Bioenerg. 2019 Aug 1;1860(8):618-627. doi: 10.1016/j.bbabio.2019.06.017. Epub 2019 Jun 25. Biochim Biophys Acta Bioenerg. 2019. PMID: 31251900
-
Clarifying the supercomplex: the higher-order organization of the mitochondrial electron transport chain.Nat Struct Mol Biol. 2017 Oct 5;24(10):800-808. doi: 10.1038/nsmb.3460. Nat Struct Mol Biol. 2017. PMID: 28981073 Review.
Cited by
-
Kingdom-specific lipid unsaturation calibrates sequence evolution in membrane arm subunits of eukaryotic respiratory complexes.Nat Commun. 2025 Feb 27;16(1):2044. doi: 10.1038/s41467-025-57295-7. Nat Commun. 2025. PMID: 40016208 Free PMC article.
-
Structures of Tetrahymena thermophila respiratory megacomplexes on the tubular mitochondrial cristae.Nat Commun. 2023 May 29;14(1):2542. doi: 10.1038/s41467-023-38158-5. Nat Commun. 2023. PMID: 37248254 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
