

Directed Evolution of Microbial Communities
- PMID: 33646814
- PMCID: PMC8105285
- DOI: 10.1146/annurev-biophys-101220-072829
Directed Evolution of Microbial Communities
Abstract
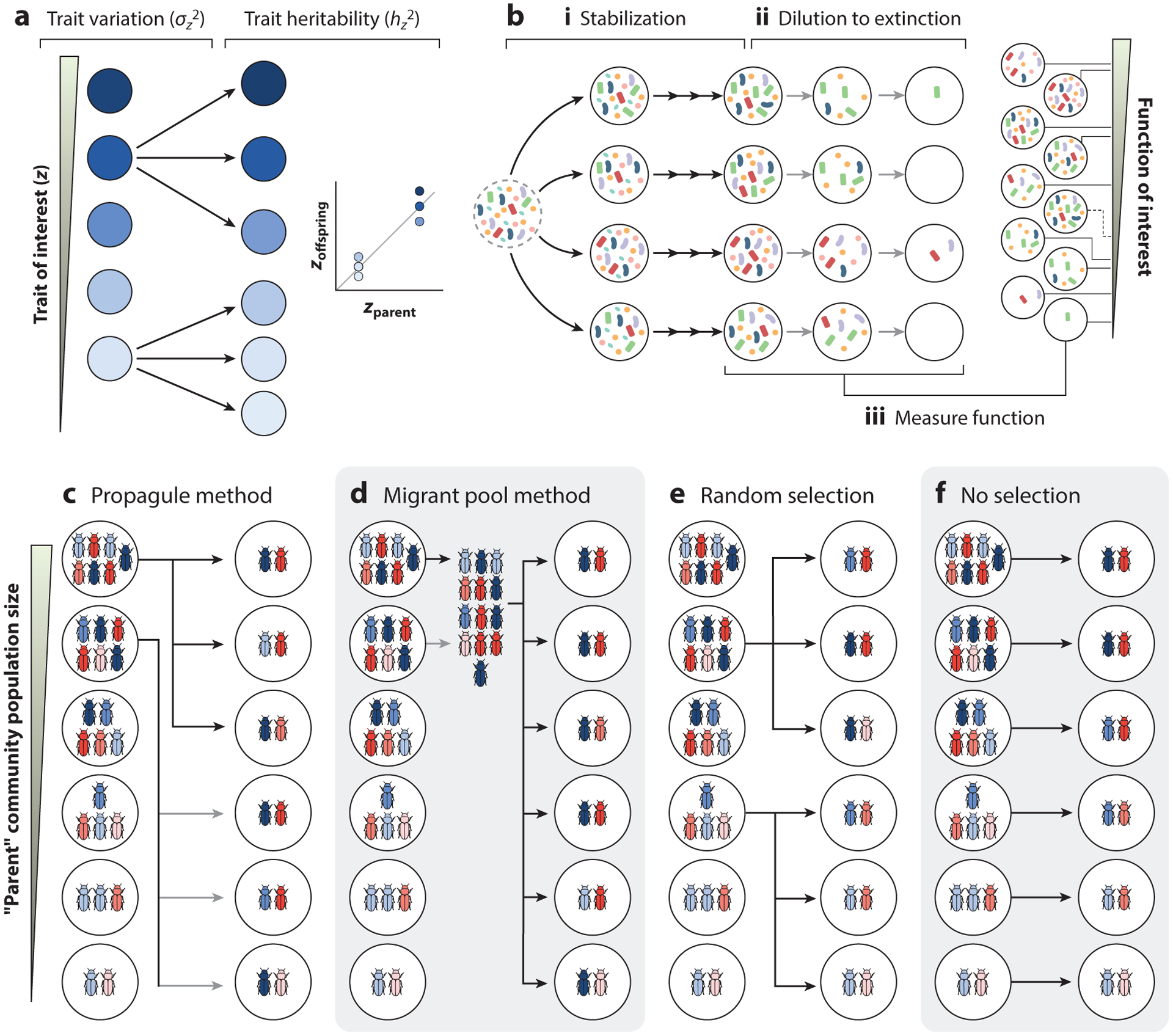
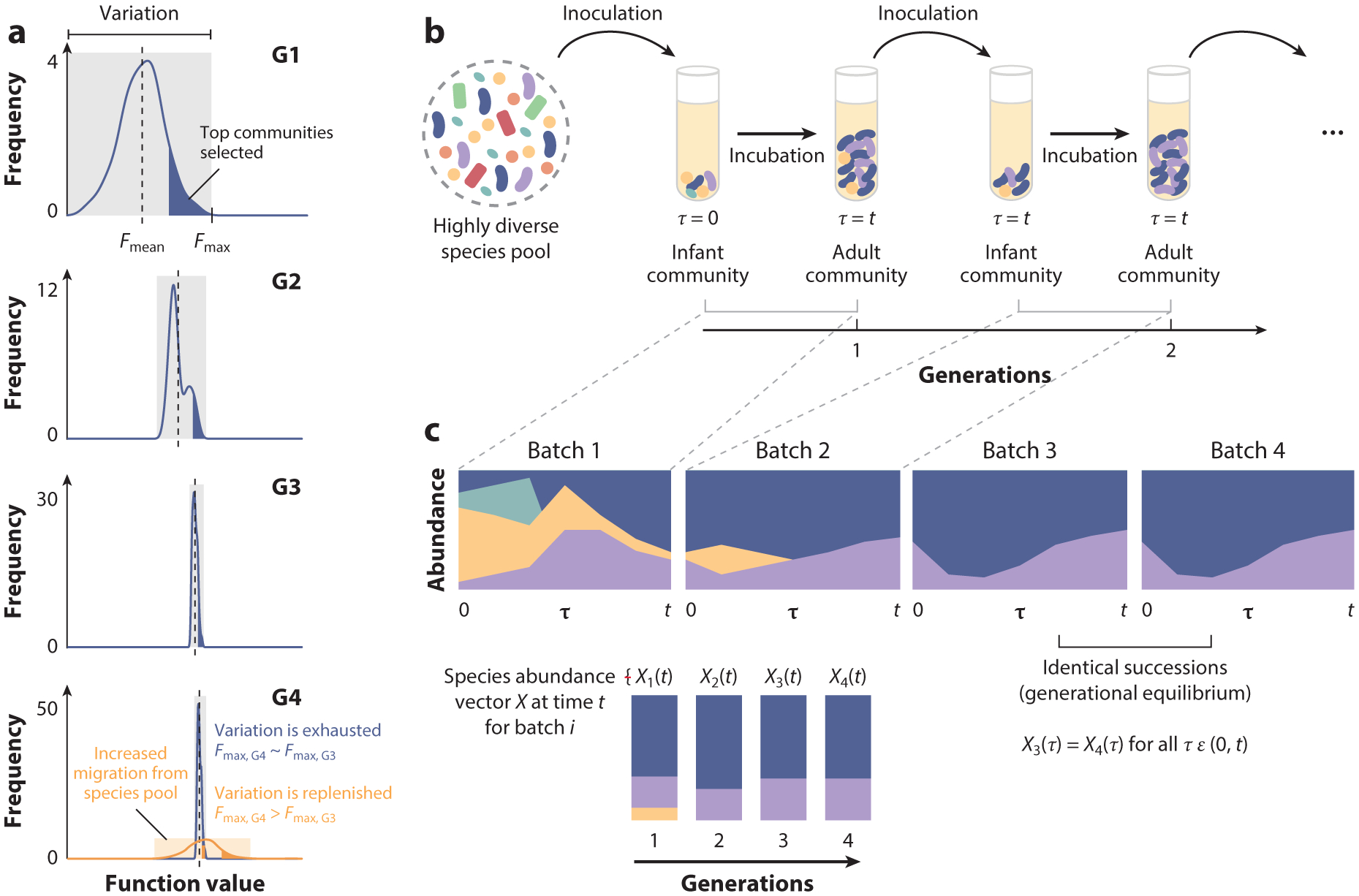
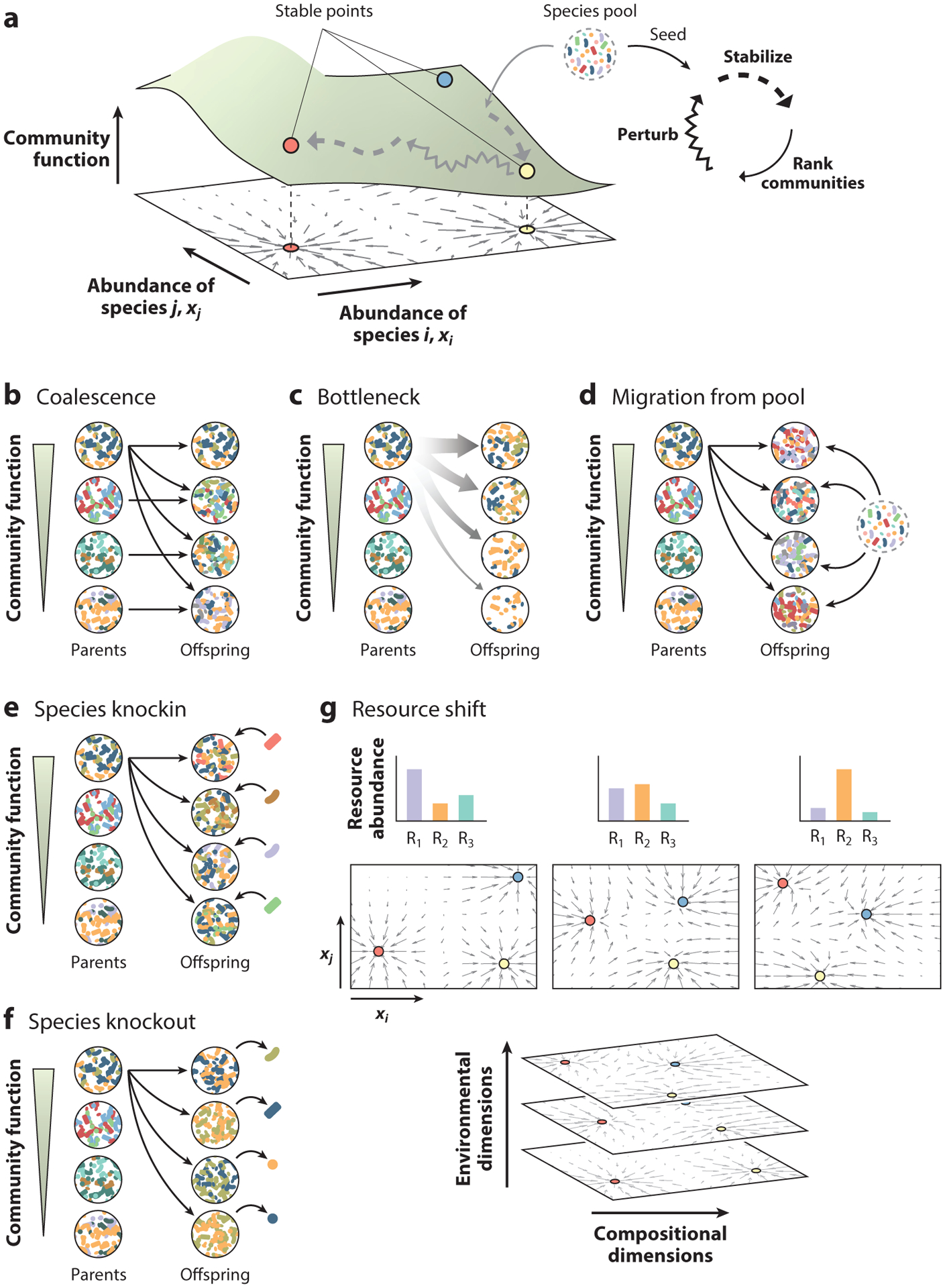
Directed evolution is a form of artificial selection that has been used for decades to find biomolecules and organisms with new or enhanced functional traits. Directed evolution can be conceptualized as a guided exploration of the genotype-phenotype map, where genetic variants with desirable phenotypes are first selected and then mutagenized to search the genotype space for an even better mutant. In recent years, the idea of applying artificial selection to microbial communities has gained momentum. In this article, we review the main limitations of artificial selection when applied to large and diverse collectives of asexually dividing microbes and discuss how the tools of directed evolution may be deployed to engineer communities from the top down. We conceptualize directed evolution of microbial communities as a guided exploration of an ecological structure-function landscape and propose practical guidelines for navigating these ecological landscapes.
Keywords: artificial ecosystem selection; collective community functions; directed evolution; microbial communities; structure–function landscape.
Figures
References
-
- Acevedo-Rocha CG, Hoebenreich S, Reetz MT. 2014. Iterative saturation mutagenesis: a powerful approach to engineer proteins by systematically simulating Darwinian evolution. In Directed Evolution Library Creation: Methods and Protocols, ed. Gillam EMJ, Copp JN, Ackerley D, pp. 103–28. Berlin: Springer - PubMed
-
- Ackermann M, Stecher B, Freed NE, Songhet P, Hardt W-D, Doebeli M. 2008. Self-destructive cooeration mediated by phenotypic noise. Nature 454(7207):987–90 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
