

Review
doi: 10.1101/cshperspect.a040543.
Mitochondria
Affiliations
- PMID: 33649187
- PMCID: PMC7919390
- DOI: 10.1101/cshperspect.a040543
Item in Clipboard
Review
Mitochondria
Cold Spring Harb Perspect Biol.
.
No abstract available
Figures
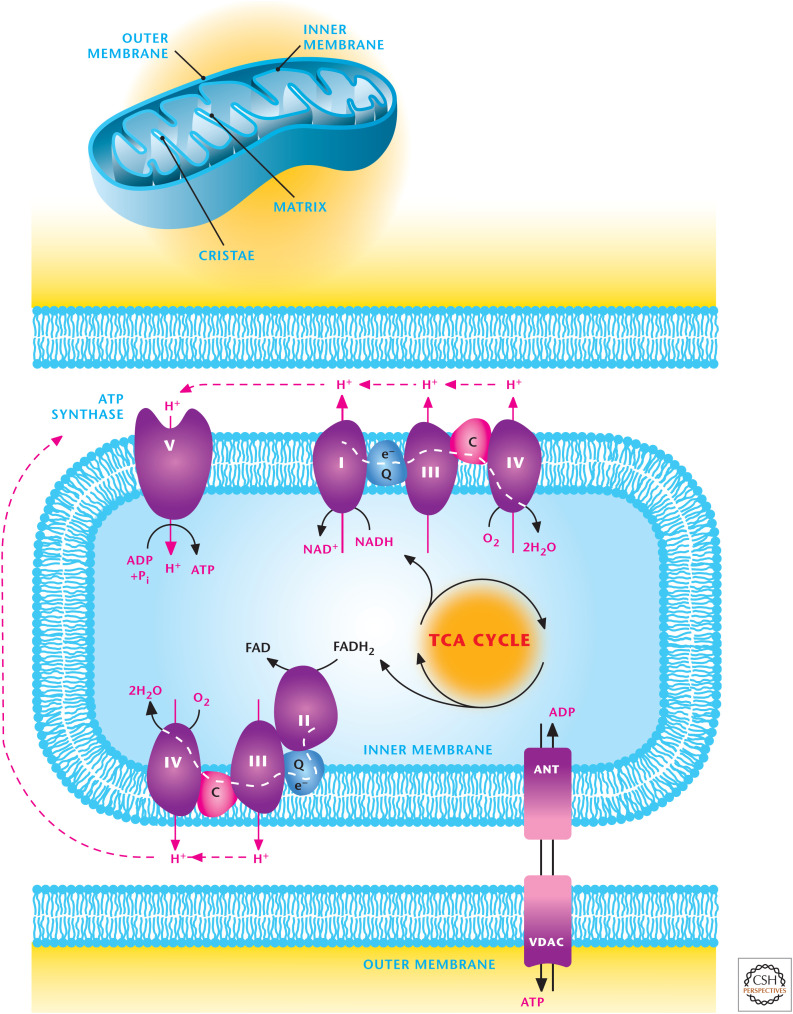
Overview of mitochondrial metabolism. Mitochondria contain an outer and inner membrane. The mitochondrial matrix enclosed by the inner membrane contains the TCA cycle that generates reducing equivalents NADH and FADH2, which donate electrons to the electron transport chain (ETC), resulting in the generation of a proton motive force to drive ATP synthesis. The ETC is composed of two supercomplexes consisting of either complex I–III–IV or II–III–IV. (Modified from http://en.wikipedia.org/wiki/Electron_transport_chain#mediaviewer/File:Mitochondrial_electron_transport_chain%E2%80%94Etc4.svg .)
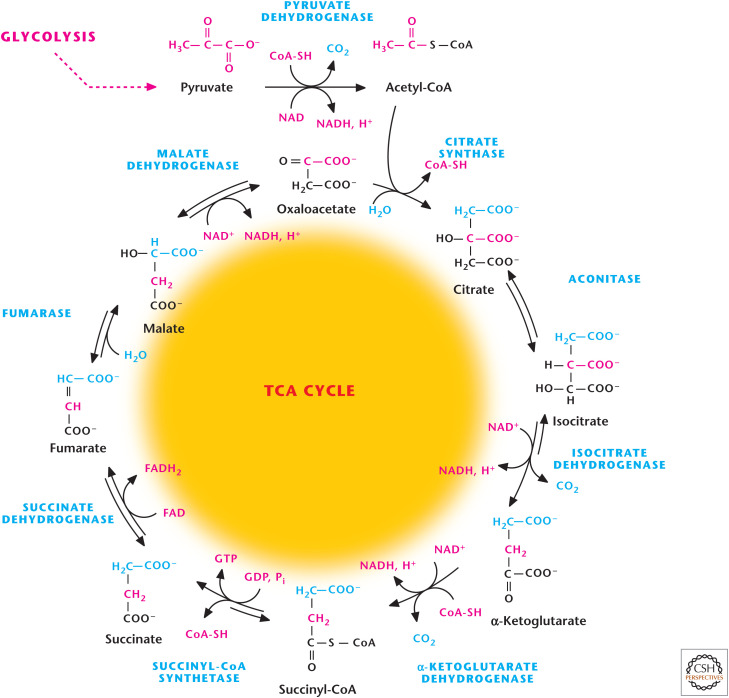
Overview of TCA cycle. The TCA cycle starts when two-carbon acetyl-CoA combine with four-carbon OAA to generate citrate. The TCA cycle produces three NADH, one FADH2, and one GTP.
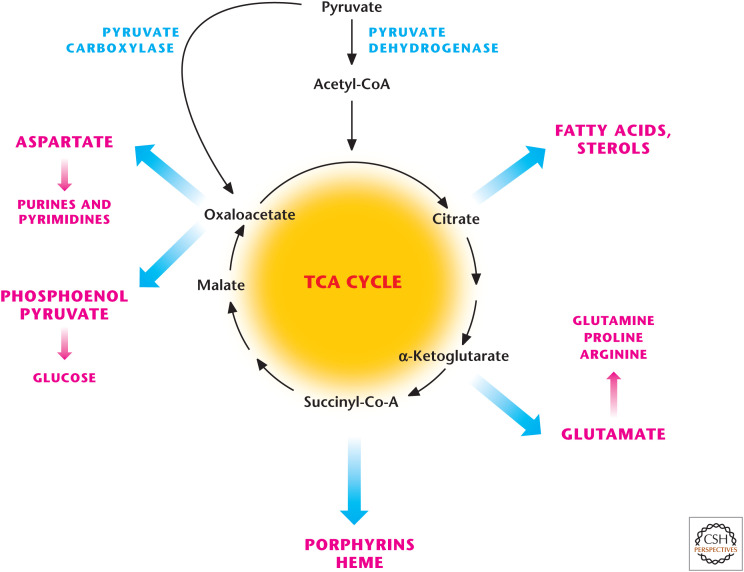
The TCA cycle is a biosynthetic hub. TCA-cycle intermediates provided the building blocks for macromolecules including fatty acids, nucleotides, hemes, and porphyrins. Oxaloacetate provides the carbons for the generation for glucose (i.e., gluconeogenesis).
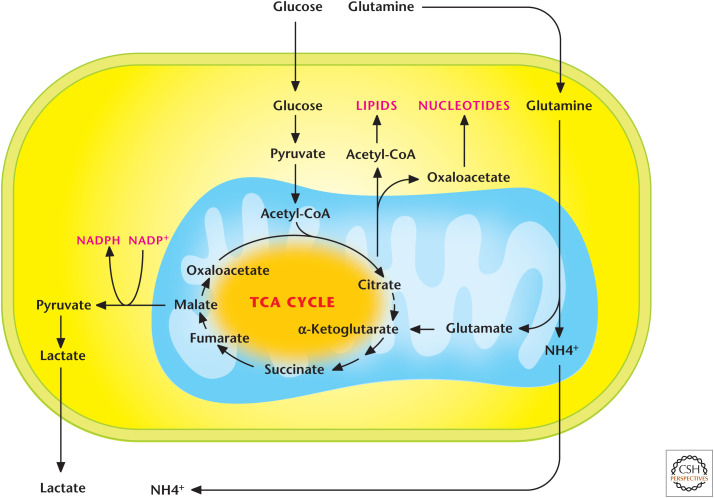
Glutaminolysis. The conversion of glutamine into glutamate and, subsequently, into α-ketoglutarate is referred to as glutaminolysis. This is an important pathway to replenish TCA-cycle intermediates.

Regulation of the TCA cycle. Citrate synthase, as well as the three dehydrogenases, isocitrate, α-ketoglutarate, and PDH, are stimulated when ADP levels increase and dampened when NADH levels increase. The three dehydrogenases are positively regulated by calcium.
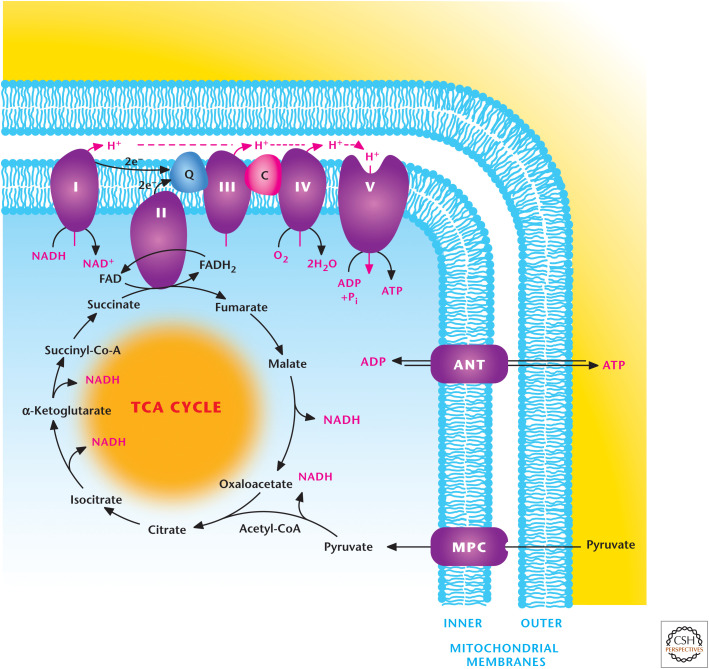
Oxidative phosphorylation. The electron transport chain complex elements I, III, and IV pump protons across the inner mitochondrial membrane as electrons are transferred through the chain to oxygen, the final electron acceptor. This protein pumping generates the pmf, composed of a small chemical component (ΔpH) and a large electrical component (membrane potential, Δψ). Complex V, the ATP synthase, uses the proton gradient established across the inner membrane to generate ATP from ADP and Pi, a process referred as oxidative phosphorylation.
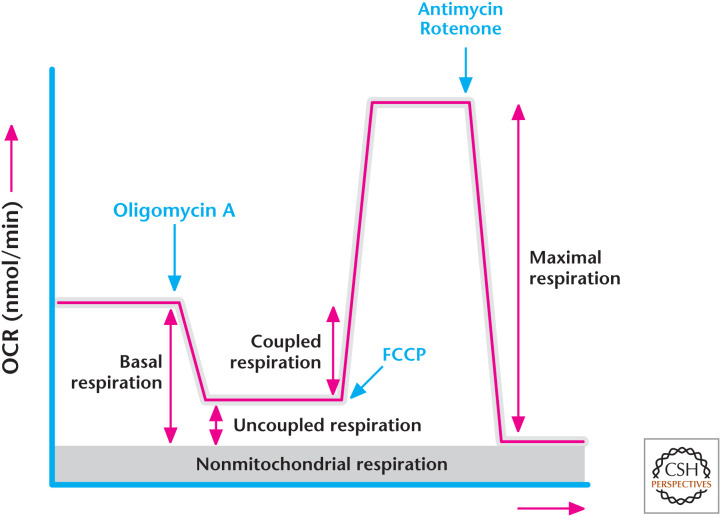
Determining the oxygen consumption rate. (Modified from Anso et al. 2013.)
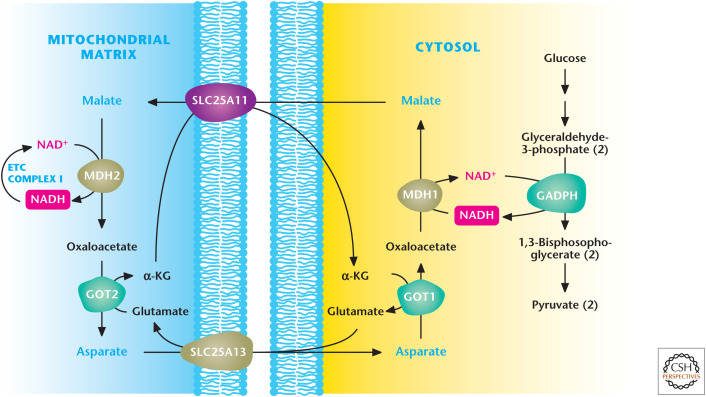
Malate–aspartate shuttle. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) generates NADH during glycolysis. NADH can be regenerated to NAD+ to allow glycolysis to continue by the conversion of oxaloacetate (OAA) to malate by cytosolic malate dehydrogenase 1 (MDH1). Subsequently, malate is transported by the SLC25A1 transporter into the mitochondrial matrix, in which it is converted back to OAA coupled with NAD+ conversion into NADH. The ETC complex I converts NADH into NAD+ to keep malate dehydrogenase 2 functioning continuously. The mitochondrial OAA is converted into aspartate by aspartate aminotransferase 2 (GOT2) and, subsequently, transported into the cytosol, where aspartate can be converted back into cytosolic OAA by aspartate aminotransferase 1 (GOT1) for the shuttle to continue. α-KG, α-ketoglutarate. (Adapted with permission of themedicalbiochemistrypage, LLC.)
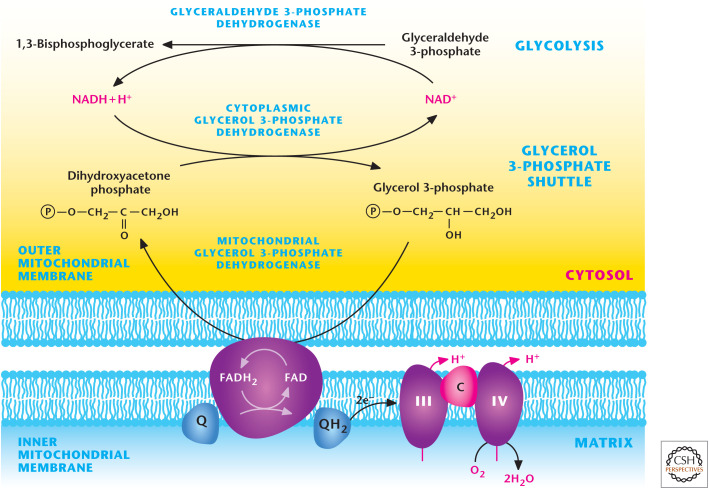
Glycerol–phosphate shuttle. GAPDH generates NADH during glycolysis. NADH can be regenerated to NAD+ to allow glycolysis to continue by the conversion of dihydroxyacetone phosphate into glycerol 3-phosphate by cytosolic glycerol 3-phosphate dehydrogenase. Subsequently, glycerol 3-phosphate is converted back into dihydroxyacetone phosphate by coupling FAD into FADH2. FADH2 donates electrons to ubiquinone, which feeds electrons into complex III of the ETC.
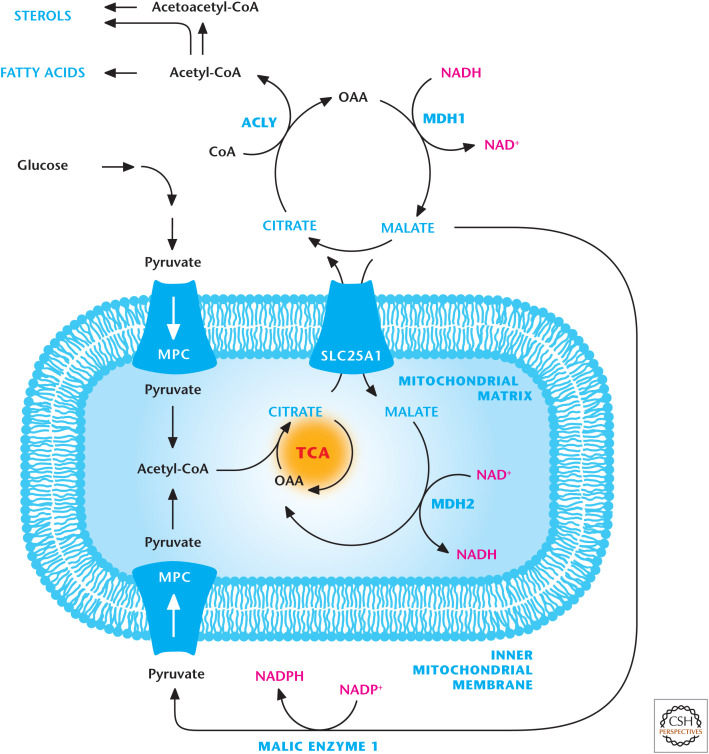
Citrate transporter. Citrate is transported into the cytosol by the SLC25A1 transporter in exchange for malate. Citrate in the cytosol is converted into acetyl-CoA and oxaloacetate (OAA) by ACLY. Acetyl-CoA is a precursor for sterol and fatty acid production. MDH1 converts the OAA into malate, which is transported into the mitochondrial matrix in exchange for citrate. Malate can also be converted into pyruvate by malic enzyme 1 (ME1).
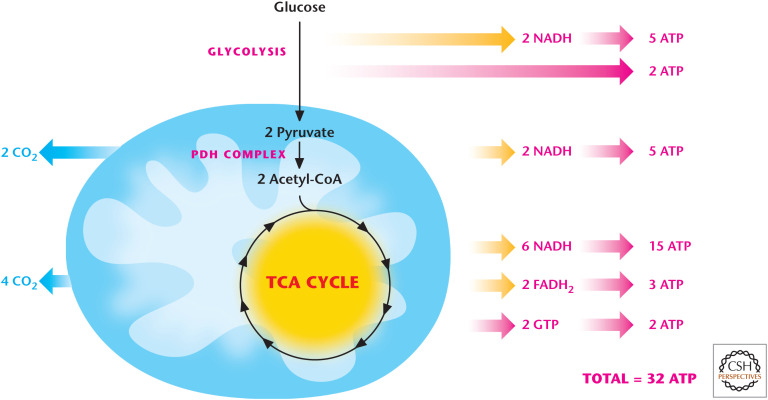
Glucose oxidation generates 32 ATP. NADH generates 2.5 ATP, whereas FADH2 generates 1.5 ATP. Glucose oxidation through glycolysis and TCA cycle generates 10 NADH (25 ATP) and 2 FADH2 (3 ATP). Glycolysis and TCA cycle generate 2 ATP each through substrate level phosphorylation.
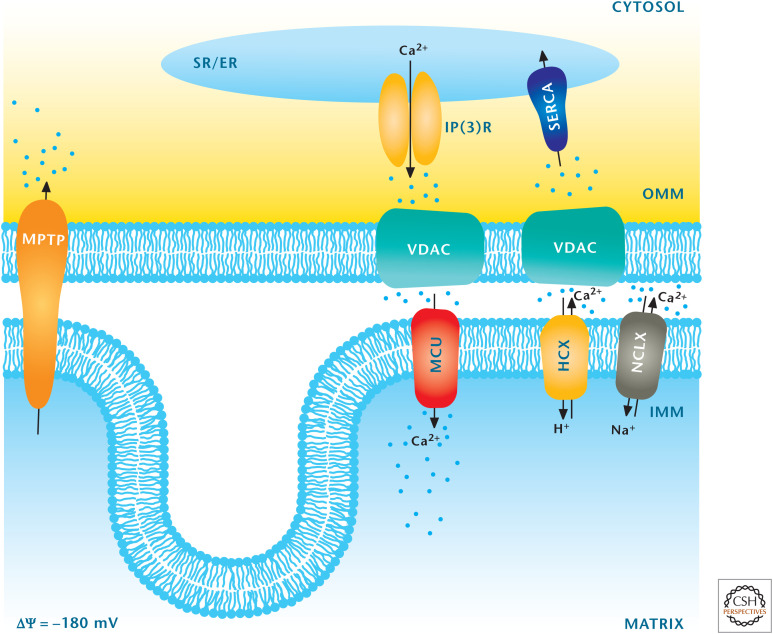
Mitochondria regulate calcium homeostasis. The major organelle that regulates calcium is the ER, which takes up calcium into the ER by sarco/ER Ca2+-ATPase (SERCA) and releases calcium from the ER into the cytosol by inositol trisphosphate receptor (InsP3R). Mitochondria can also regulate calcium by sequestering cytosolic calcium through the MCU. Calcium is transported from mitochondria into the cytosol through the mitochondrial H+–Ca2+ exchanger (HCX) and mitochondrial Na+–Ca2+ exchanger (NCLX) transporters. (Adapted from Raffaello et al. 2012, with permission from Elsevier.)
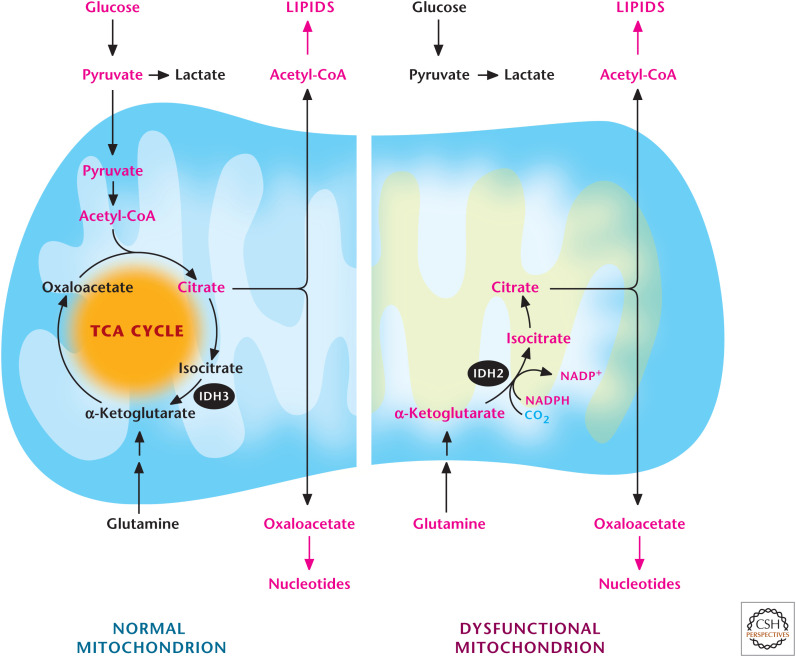
Glutamine-dependent reductive carboxylation. Cells that have functional mitochondria can use glutamine to generate α-ketoglutarate to produce OAA and use pyruvate to generate acetyl-CoA. OAA and acetyl-CoA generate citrate, which can be exported into the cytosol to produce de novo lipids and nucleotides. Cells that have dysfunctional mitochondria due to loss-of-function mutations of proteins in the TCA cycle after the α-ketoglutarate dehydrogenase step or in the ETC cannot oxidize pyruvate into acetyl-CoA. They convert glutamine into α-ketoglutarate, which subsequently becomes citrate through a reverse IDH2-dependent reaction.
References
-
- Chance B, Williams GR. 1955. Respiratory enzymes in oxidative phosphorylation. III. The steady state. J Biol Chem 217: 409–427. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources