

Lipid droplets in mammalian eggs are utilized during embryonic diapause
- PMID: 33649221
- PMCID: PMC7958255
- DOI: 10.1073/pnas.2018362118
Lipid droplets in mammalian eggs are utilized during embryonic diapause
Abstract
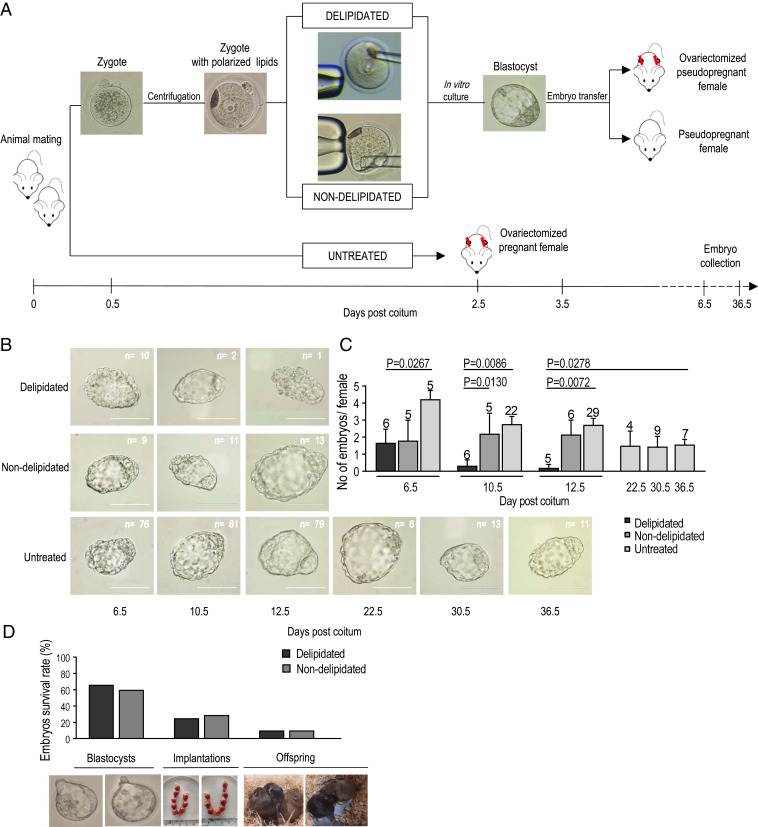
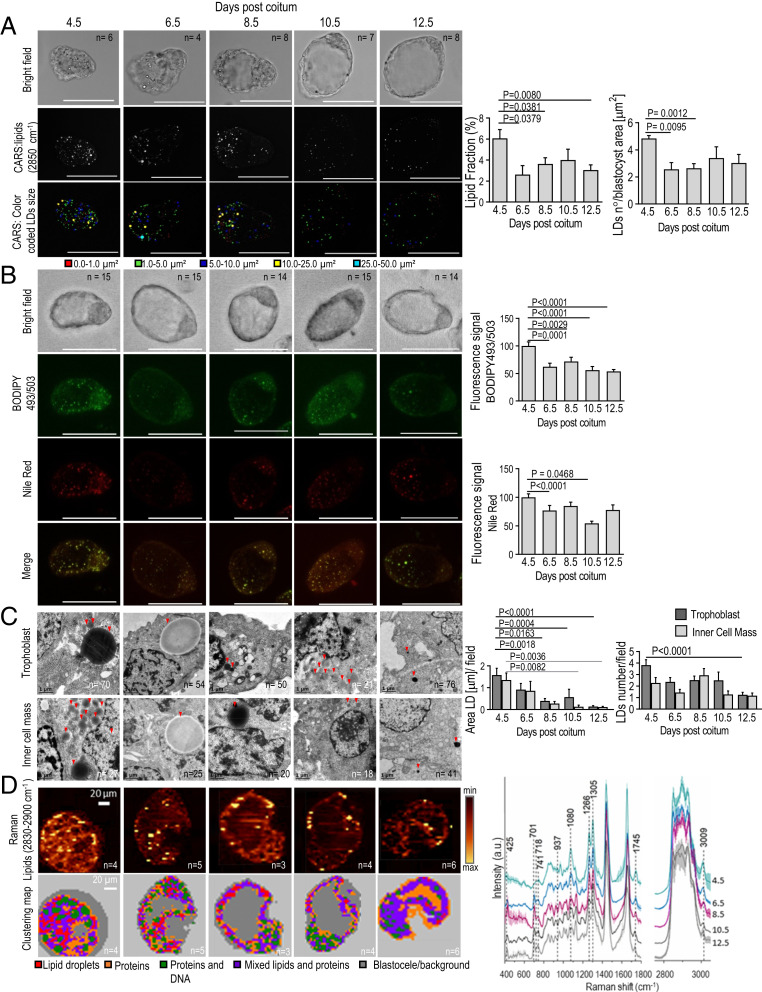
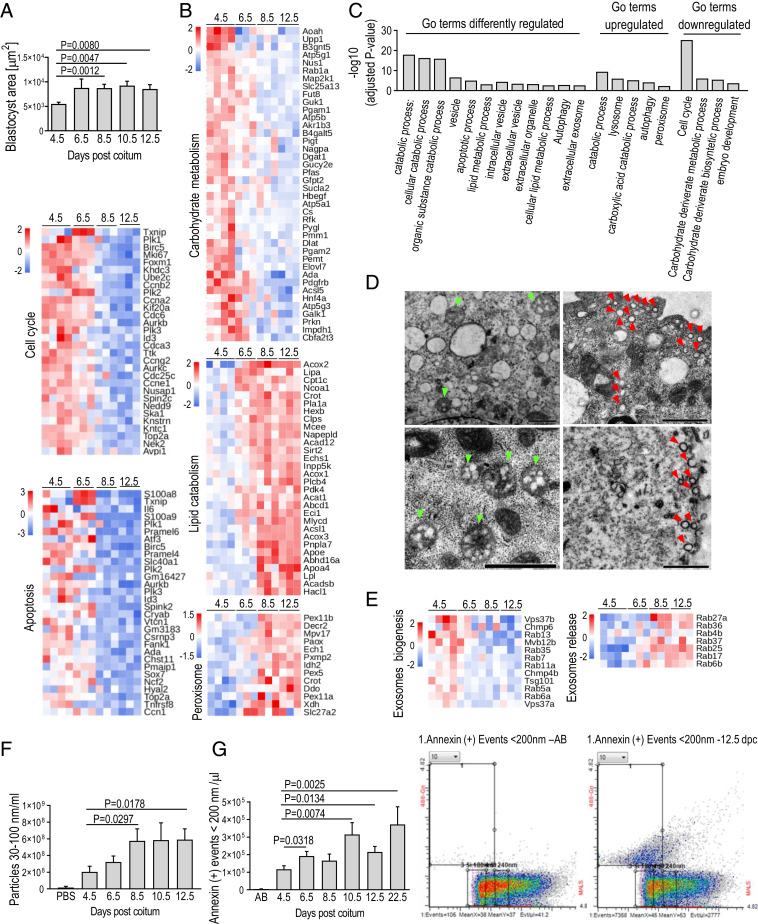
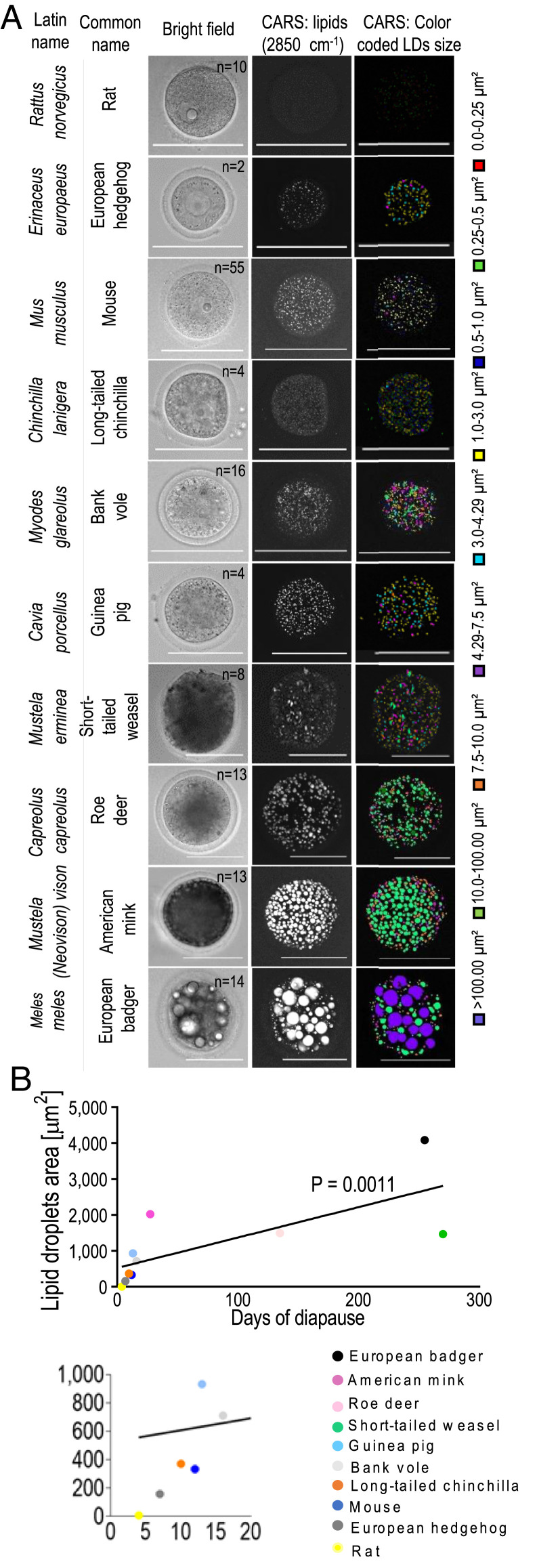
Embryonic diapause (ED) is a temporary arrest of an embryo at the blastocyst stage when it waits for the uterine receptivity signal to implant. ED used by over 100 species may also occur in normally "nondiapausing" mammals when the uterine receptivity signal is blocked or delayed. A large number of lipid droplets (LDs) are stored throughout the preimplantation embryo development, but the amount of lipids varies greatly across different mammalian species. Yet, the role of LDs in the mammalian egg and embryo remains unknown. Here, using a mouse model, we provide evidence that LDs play a crucial role in maintaining ED. By mechanical removal of LDs from zygotes, we demonstrated that delipidated embryos are unable to survive during ED. LDs are not essential for normal prompt implantation, without ED. We further demonstrated that with the progression of ED, the amount of intracellular lipid reduces, and composition changes. This decrease in lipid is caused by a switch from carbohydrate metabolism to lipid catabolism in diapausing blastocysts, which also exhibit increased release of exosomes reflecting elevated embryonic signaling to the mother. We have also shown that presence of LDs in the oocytes of various mammals positively corelates with their species-specific length of diapause. Our results reveal the functional role of LDs in embryonic development. These results can help to develop diagnostic techniques and treatment of recurrent implantation failure and will likely ignite further studies in developmental biology and reproductive medicine fields.
Keywords: blastocyst; embryonic diapause; lipid droplets.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Barbosa A. D., Savage D. B., Siniossoglou S., Lipid droplet-organelle interactions: Emerging roles in lipid metabolism. Curr. Opin. Cell Biol. 35, 91–97 (2015). - PubMed
-
- Nagashima H., Kashiwazaki N., Ashman R. J., Grupen C. G., Nottle M. B., Cryopreservation of porcine embryos. Nature 374, 416 (1995). - PubMed
-
- Renfree M. B., Fenelon J. C., The enigma of embryonic diapause. Development 144, 3199–3210 (2017). - PubMed
-
- Chang M. C., Reciprocal insemination and egg transfer between ferrets and mink. J. Exp. Zool. 168, 49–59 (1968). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
