

Deubiquitinase USP39 and E3 ligase TRIM26 balance the level of ZEB1 ubiquitination and thereby determine the progression of hepatocellular carcinoma
- PMID: 33649471
- PMCID: PMC8329202
- DOI: 10.1038/s41418-021-00754-7
Deubiquitinase USP39 and E3 ligase TRIM26 balance the level of ZEB1 ubiquitination and thereby determine the progression of hepatocellular carcinoma
Abstract
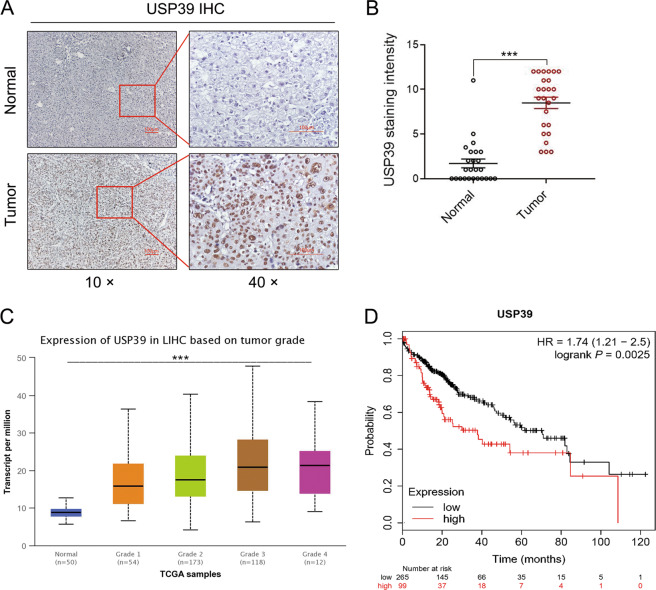
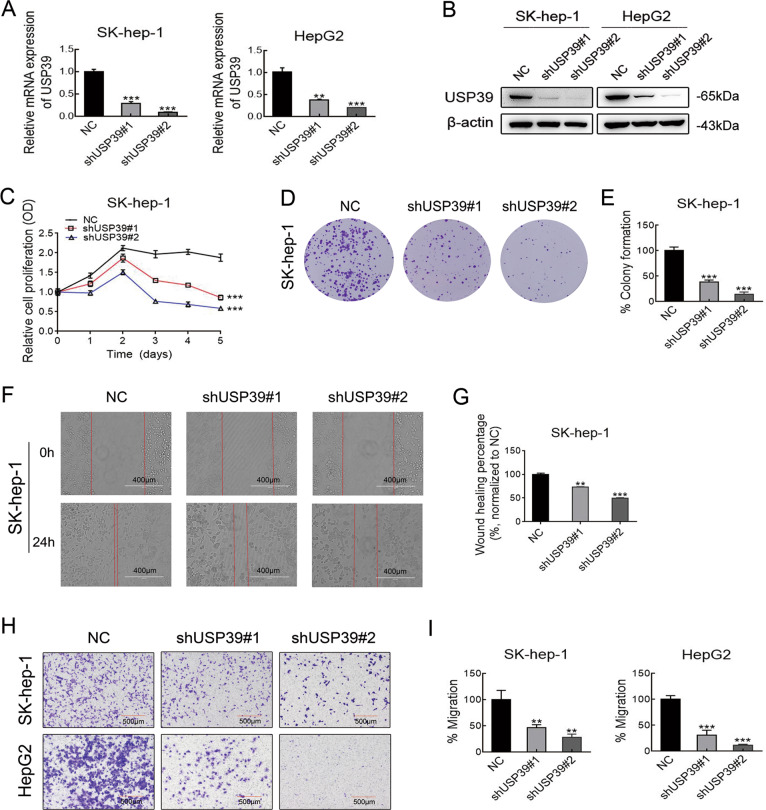
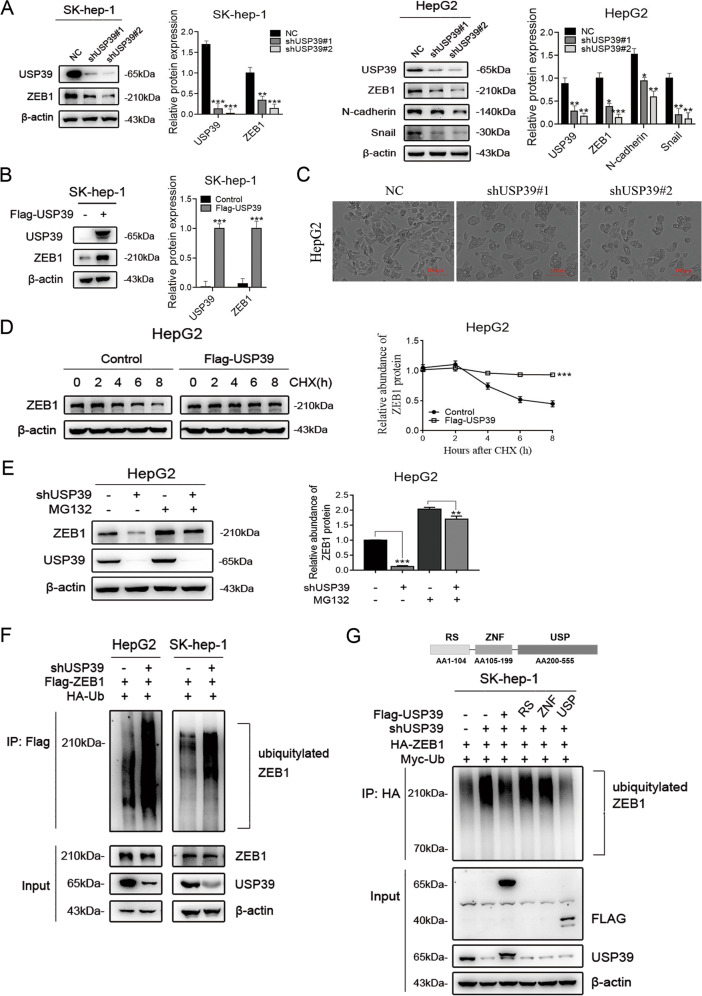
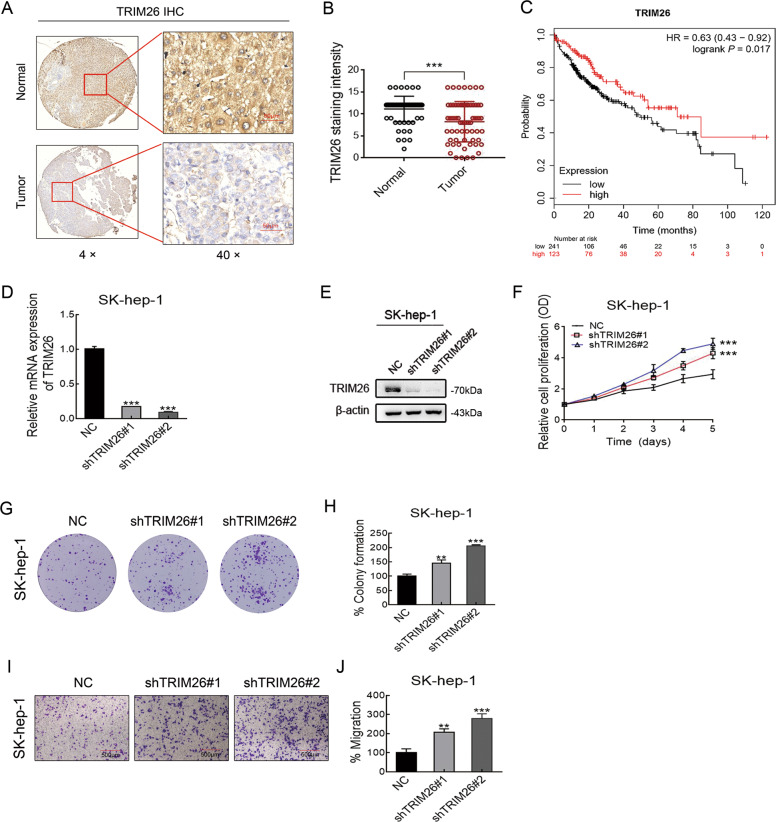
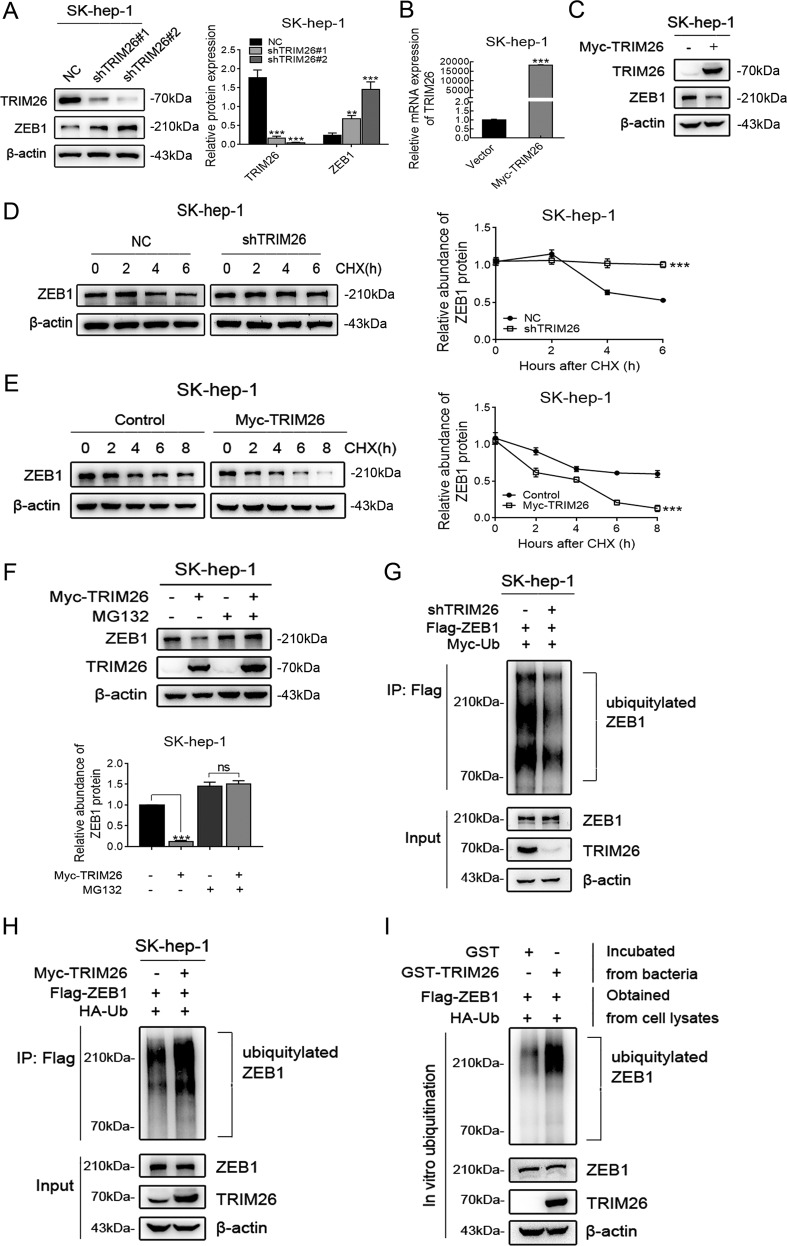
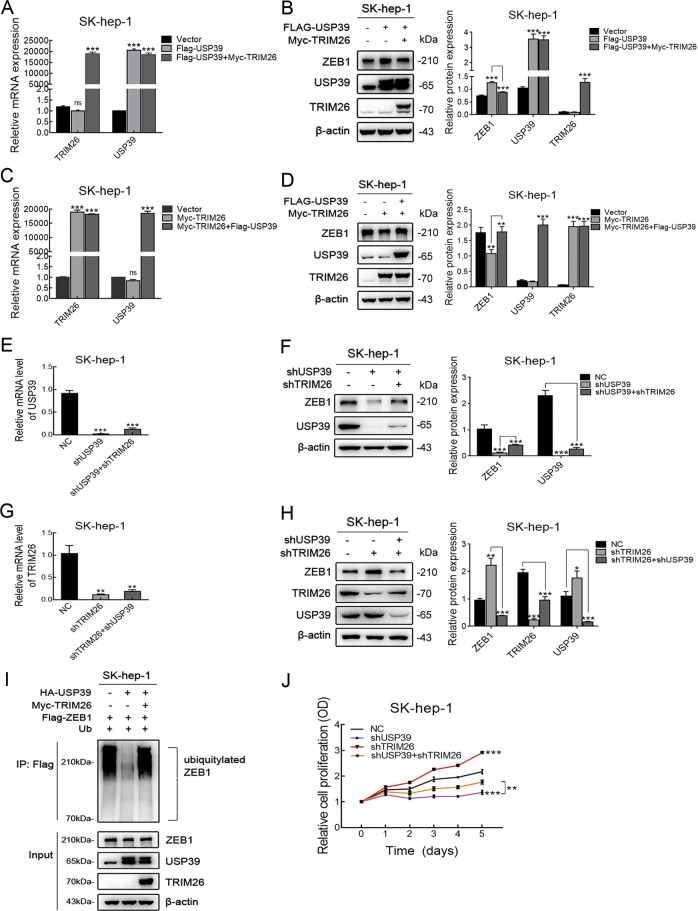
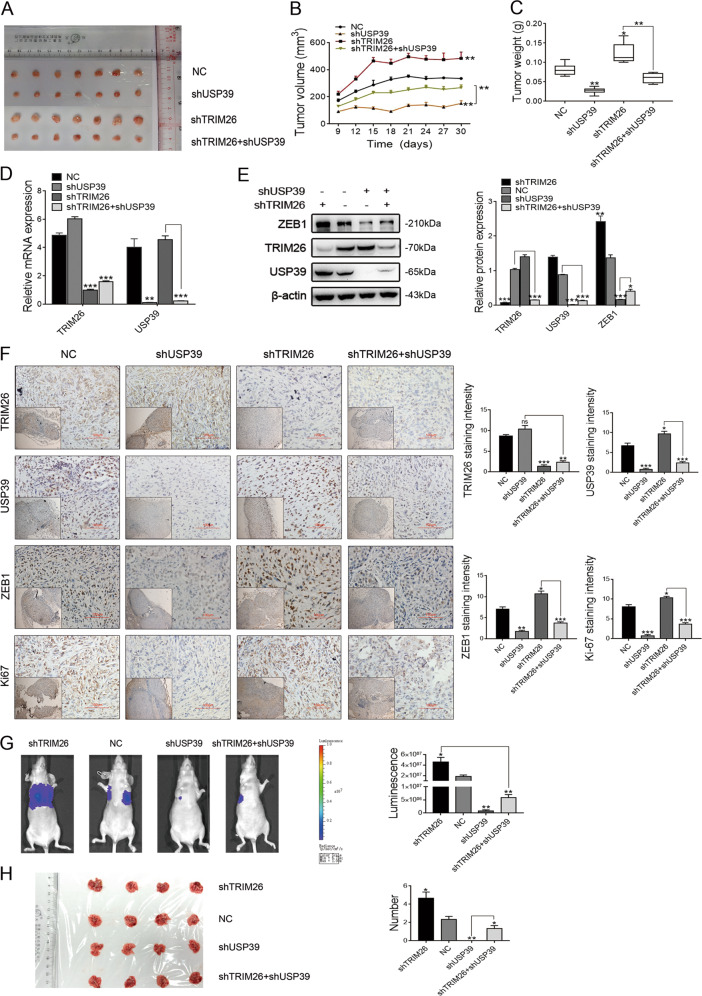
Emerging evidence suggests that USP39 plays an important role in the development of hepatocellular carcinoma (HCC). However, the molecular mechanism by which USP39 promotes HCC progression has not been well defined, especially regarding its putative ubiquitination function. Zinc-finger E-box-binding homeobox 1 (ZEB1) is a crucial inducer of epithelial-to-mesenchymal transition (EMT) to promote tumor proliferation and metastasis, but the regulatory mechanism of ZEB1 stability in HCC remains enigmatic. Here, we reveal that USP39 is highly expressed in human HCC tissues and correlated with poor prognosis. Moreover, USP39 depletion inhibits HCC cell proliferation and metastasis by promoting ZEB1 degradation. Intriguingly, deubiquitinase USP39 has a direct interaction with the E3 ligase TRIM26 identified by co-immunoprecipitation assays and immunofluorescence staining assays. We further demonstrate that TRIM26 is lowly expressed in human HCC tissues and inhibits HCC cell proliferation and migration. TRIM26 promotes the degradation of ZEB1 protein by ubiquitination in HCC. Deubiquitinase USP39 and E3 ligase TRIM26 function in an antagonistic pattern, but not a competitive pattern, and play key roles in controlling ZEB1 stability to determine the HCC progression. In summary, our data reveal a previously unknown mechanism that USP39 and TRIM26 balance the level of ZEB1 ubiquitination and thereby determine HCC cell proliferation and migration. This novel mechanism may provide new approaches to target treatment for inhibiting HCC development by restoring TRIM26 or suppressing USP39 expression in HCC cases with high ZEB1 protein levels.
© 2021. The Author(s), under exclusive licence to ADMC Associazione Differenziamento e Morte Cellulare.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
