

Escherichia coli segments its controls on carbon-dependent gene expression into global and specific regulations
- PMID: 33650807
- PMCID: PMC8085971
- DOI: 10.1111/1751-7915.13776
Escherichia coli segments its controls on carbon-dependent gene expression into global and specific regulations
Abstract
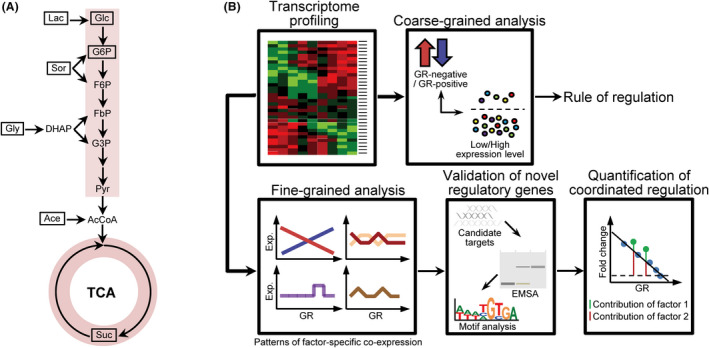
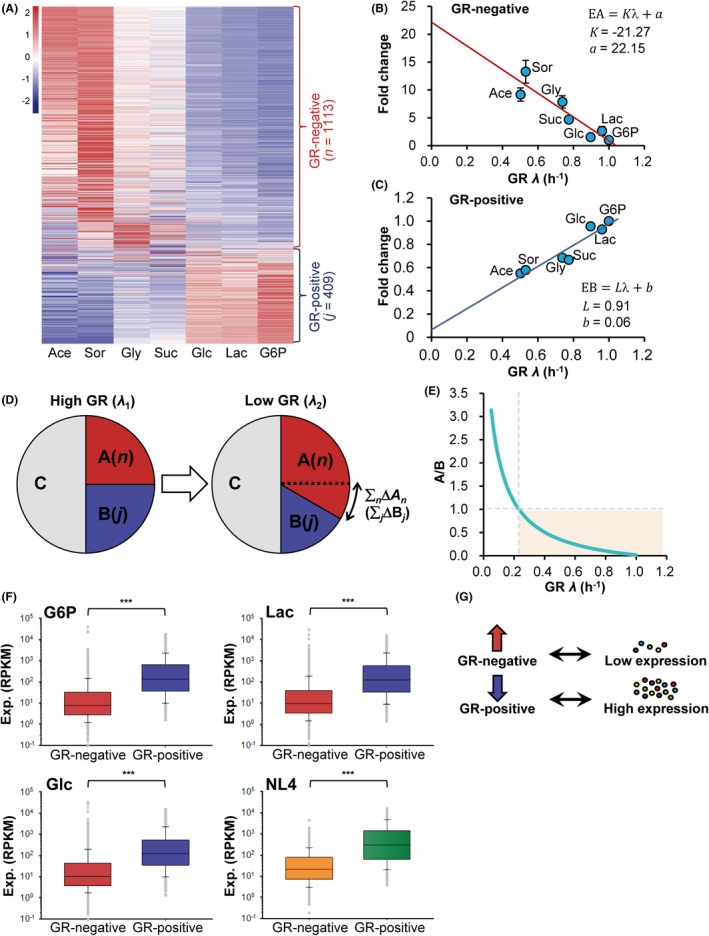
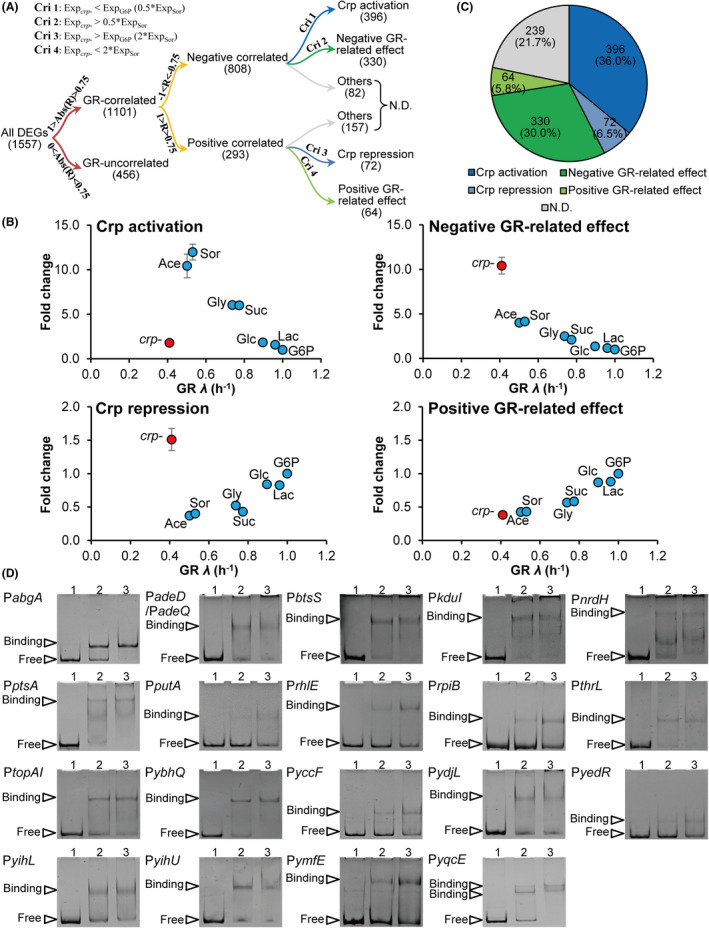
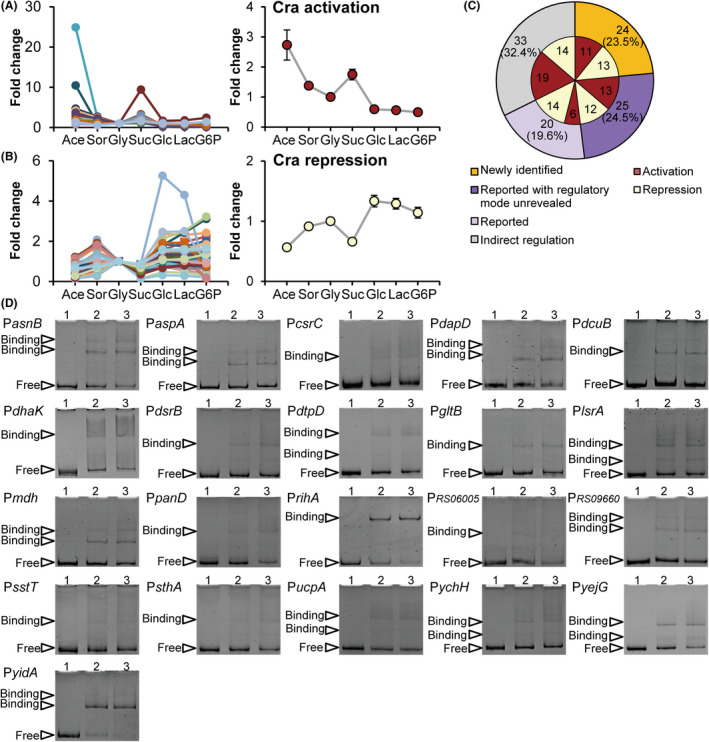
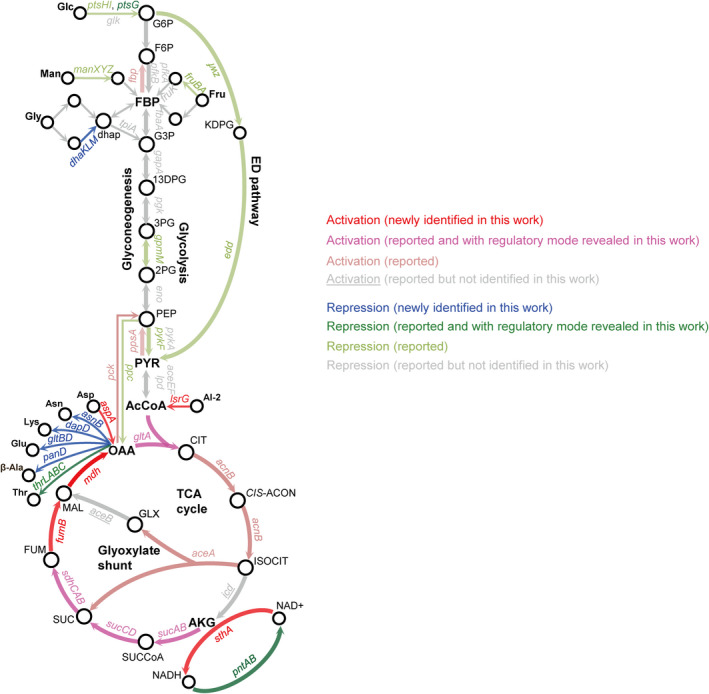
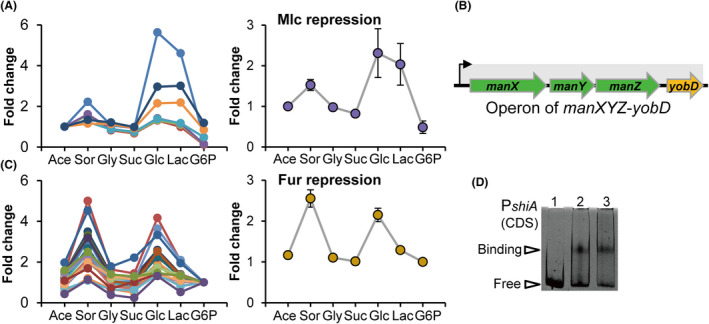
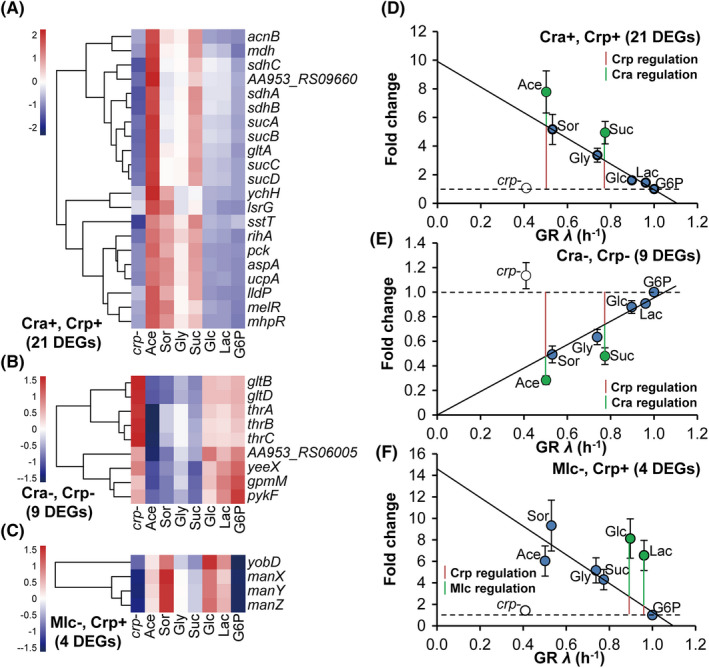
How bacteria adjust gene expression to cope with variable environments remains open to question. Here, we investigated the way global gene expression changes in E. coli correlated with the metabolism of seven carbon substrates chosen to trigger a large panel of metabolic pathways. Coarse-grained analysis of gene co-expression identified a novel regulation pattern: we established that the gene expression trend following immediately the reduction of growth rate (GR) was correlated to its initial expression level. Subsequent fine-grained analysis of co-expression demonstrated that the Crp regulator, coupled with a change in GR, governed the response of most GR-dependent genes. By contrast, the Cra, Mlc and Fur regulators governed the expression of genes responding to non-glycolytic substrates, glycolytic substrates or phosphotransferase system transported sugars following an idiosyncratic way. This work allowed us to expand additional genes in the panel of gene complement regulated by each regulator and to elucidate the regulatory functions of each regulator comprehensively. Interestingly, the bulk of genes controlled by Cra and Mlc were, respectively, co-regulated by Crp- or GR-related effect and our quantitative analysis showed that each factor took turns to work as the primary one or contributed equally depending on the conditions.
© 2021 The Authors. Microbial Biotechnology published by John Wiley & Sons Ltd and Society for Applied Microbiology.
Conflict of interest statement
None declared.
Figures
Similar articles
-
Transcriptional regulation of main metabolic pathways of cyoA, cydB, fnr, and fur gene knockout Escherichia coli in C-limited and N-limited aerobic continuous cultures.Microb Cell Fact. 2011 Jan 27;10:3. doi: 10.1186/1475-2859-10-3. Microb Cell Fact. 2011. PMID: 21272324 Free PMC article.
-
Functional interactions between the carbon and iron utilization regulators, Crp and Fur, in Escherichia coli.J Bacteriol. 2005 Feb;187(3):980-90. doi: 10.1128/JB.187.3.980-990.2005. J Bacteriol. 2005. PMID: 15659676 Free PMC article.
-
Microbial Cell Factories à la Carte: Elimination of Global Regulators Cra and ArcA Generates Metabolic Backgrounds Suitable for the Synthesis of Bioproducts in Escherichia coli.Appl Environ Microbiol. 2018 Sep 17;84(19):e01337-18. doi: 10.1128/AEM.01337-18. Print 2018 Oct 1. Appl Environ Microbiol. 2018. PMID: 30030227 Free PMC article.
-
Regulation of gene expression in the PTS in Escherichia coli: the role and interactions of Mlc.Curr Opin Microbiol. 2002 Apr;5(2):187-93. doi: 10.1016/s1369-5274(02)00296-5. Curr Opin Microbiol. 2002. PMID: 11934616 Review.
-
Members of the Fur protein family regulate iron and zinc transport in E. coli and characteristics of the Fur-regulated fhuF protein.J Mol Microbiol Biotechnol. 2002 May;4(3):217-22. J Mol Microbiol Biotechnol. 2002. PMID: 11931550 Review.
Cited by
-
A Systems Biology Approach To Disentangle the Direct and Indirect Effects of Global Transcription Factors on Gene Expression in Escherichia coli.Microbiol Spectr. 2023 Feb 7;11(2):e0210122. doi: 10.1128/spectrum.02101-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36749045 Free PMC article.
-
Insights into the regulatory mechanisms and application prospects of the transcription factor Cra.Appl Environ Microbiol. 2024 Nov 20;90(11):e0122824. doi: 10.1128/aem.01228-24. Epub 2024 Nov 4. Appl Environ Microbiol. 2024. PMID: 39494897 Free PMC article. Review.
-
Improving the Annotations of JCVI-Syn3a Proteins.Methods Mol Biol. 2025;2867:153-168. doi: 10.1007/978-1-0716-4196-5_9. Methods Mol Biol. 2025. PMID: 39576580 Free PMC article.
-
Patterns of Fitness and Gene Expression Epistasis Generated by Beneficial Mutations in the rho and rpoB Genes of Escherichia coli during High-Temperature Adaptation.Mol Biol Evol. 2024 Sep 4;41(9):msae187. doi: 10.1093/molbev/msae187. Mol Biol Evol. 2024. PMID: 39235107 Free PMC article.
-
Small RNA GcvB Regulates Oxidative Stress Response of Escherichia coli.Antioxidants (Basel). 2021 Nov 5;10(11):1774. doi: 10.3390/antiox10111774. Antioxidants (Basel). 2021. PMID: 34829644 Free PMC article.
References
-
- Benning, C. (1998) Biosynthesis and function of the sulfolipid sulfoquinovosyl diacylglycerol. Annu Rev Plant Physiol Plant Mol Biol 49: 53–75. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
