

Protein methyltransferase 7 deficiency in Leishmania major increases neutrophil associated pathology in murine model
- PMID: 33651805
- PMCID: PMC7954300
- DOI: 10.1371/journal.pntd.0009230
Protein methyltransferase 7 deficiency in Leishmania major increases neutrophil associated pathology in murine model
Abstract
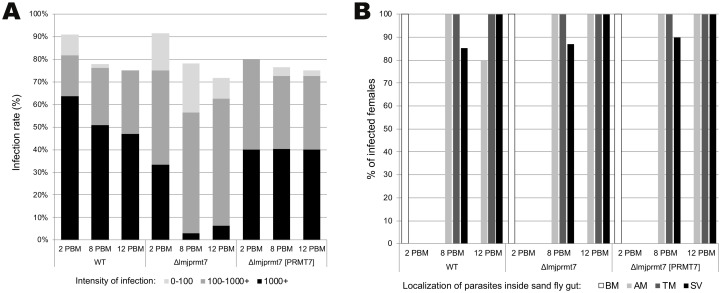
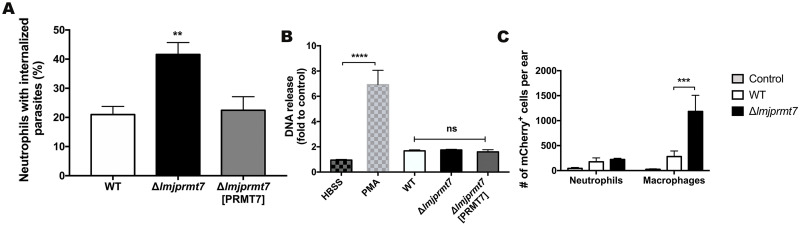
Leishmania major is the main causative agent of cutaneous leishmaniasis in the Old World. In Leishmania parasites, the lack of transcriptional control is mostly compensated by post-transcriptional mechanisms. Methylation of arginine is a conserved post-translational modification executed by Protein Arginine Methyltransferase (PRMTs). The genome from L. major encodes five PRMT homologs, including the cytosolic protein associated with several RNA-binding proteins, LmjPRMT7. It has been previously reported that LmjPRMT7 could impact parasite infectivity. In addition, a more recent work has clearly shown the importance of LmjPRMT7 in RNA-binding capacity and protein stability of methylation targets, demonstrating the role of this enzyme as an important epigenetic regulator of mRNA metabolism. In this study, we unveil the impact of PRMT7-mediated methylation on parasite development and virulence. Our data reveals that higher levels of LmjPRMT7 can impair parasite pathogenicity, and that deletion of this enzyme rescues the pathogenic phenotype of an attenuated strain of L. major. Interestingly, lesion formation caused by LmjPRMT7 knockout parasites is associated with an exacerbated inflammatory reaction in the tissue correlated with an excessive neutrophil recruitment. Moreover, the absence of LmjPRMT7 also impairs parasite development within the sand fly vector Phlebotomus duboscqi. Finally, a transcriptome analysis shed light onto possible genes affected by depletion of this enzyme. Taken together, this study highlights how post-transcriptional regulation can affect different aspects of the parasite biology.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
Altered expression of an RBP-associated arginine methyltransferase 7 in Leishmania major affects parasite infection.Mol Microbiol. 2014 Oct 8. doi: 10.1111/mmi.12819. Online ahead of print. Mol Microbiol. 2014. PMID: 25294169
-
PRMT7 regulates RNA-binding capacity and protein stability in Leishmania parasites.Nucleic Acids Res. 2020 Jun 4;48(10):5511-5526. doi: 10.1093/nar/gkaa211. Nucleic Acids Res. 2020. PMID: 32365184 Free PMC article.
-
Promastigote secretory gel from natural and unnatural sand fly vectors exacerbate Leishmania major and Leishmania tropica cutaneous leishmaniasis in mice.Parasitology. 2019 Dec;146(14):1796-1802. doi: 10.1017/S0031182019001069. Epub 2019 Aug 29. Parasitology. 2019. PMID: 31452467 Free PMC article.
-
The Impact of Neutrophil Recruitment to the Skin on the Pathology Induced by Leishmania Infection.Front Immunol. 2021 Mar 1;12:649348. doi: 10.3389/fimmu.2021.649348. eCollection 2021. Front Immunol. 2021. PMID: 33732265 Free PMC article. Review.
-
Distribution of Leishmania major zymodemes in relation to populations of Phlebotomus papatasi sand flies.Parasit Vectors. 2011 Jan 25;4:9. doi: 10.1186/1756-3305-4-9. Parasit Vectors. 2011. PMID: 21266079 Free PMC article. Review.
Cited by
-
Arginine Methyltransferases as Regulators of RNA-Binding Protein Activities in Pathogenic Kinetoplastids.Front Mol Biosci. 2021 Jun 11;8:692668. doi: 10.3389/fmolb.2021.692668. eCollection 2021. Front Mol Biosci. 2021. PMID: 34179098 Free PMC article. Review.
-
Next-Generation Leishmanization: Revisiting Molecular Targets for Selecting Genetically Engineered Live-Attenuated Leishmania.Microorganisms. 2023 Apr 16;11(4):1043. doi: 10.3390/microorganisms11041043. Microorganisms. 2023. PMID: 37110466 Free PMC article. Review.
-
High-speed, three-dimensional imaging reveals chemotactic behaviour specific to human-infective Leishmania parasites.Elife. 2021 Jun 28;10:e65051. doi: 10.7554/eLife.65051. Elife. 2021. PMID: 34180835 Free PMC article.
-
Protein arginine methyltransferases in protozoan parasites.Parasitology. 2022 Apr;149(4):427-435. doi: 10.1017/S0031182021002043. Epub 2021 Dec 6. Parasitology. 2022. PMID: 35331350 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
