

The Atypical Kinase RIOK3 Limits RVFV Propagation and Is Regulated by Alternative Splicing
- PMID: 33652597
- PMCID: PMC7996929
- DOI: 10.3390/v13030367
The Atypical Kinase RIOK3 Limits RVFV Propagation and Is Regulated by Alternative Splicing
Abstract
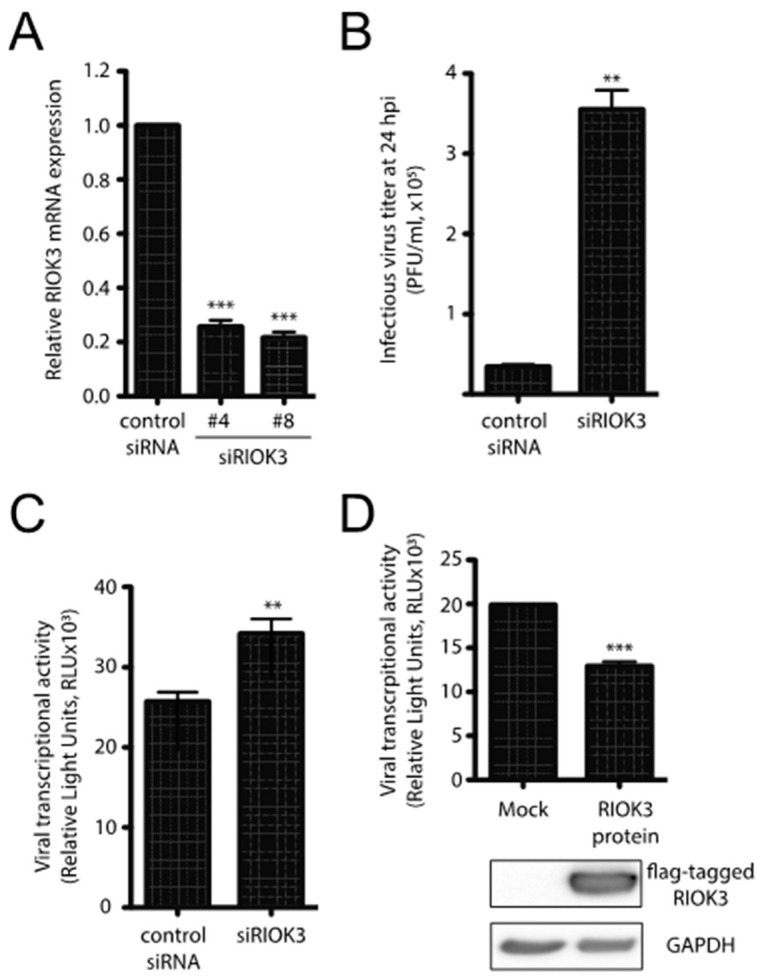
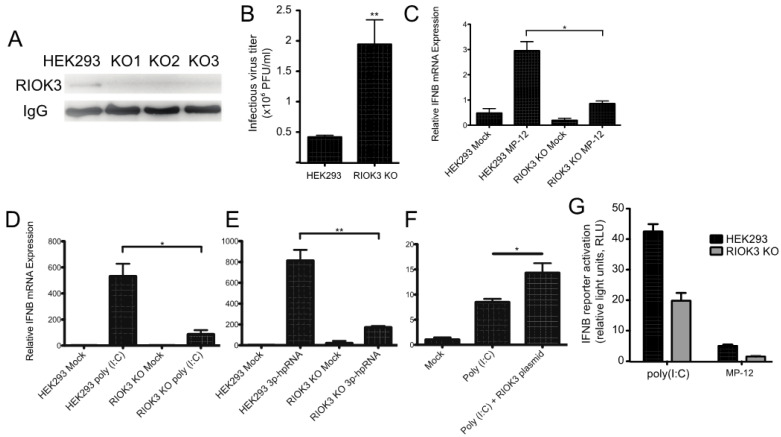
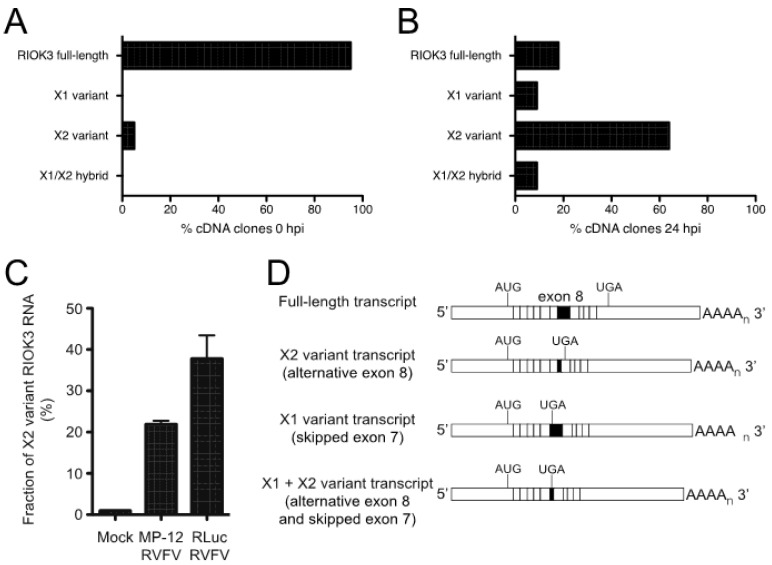
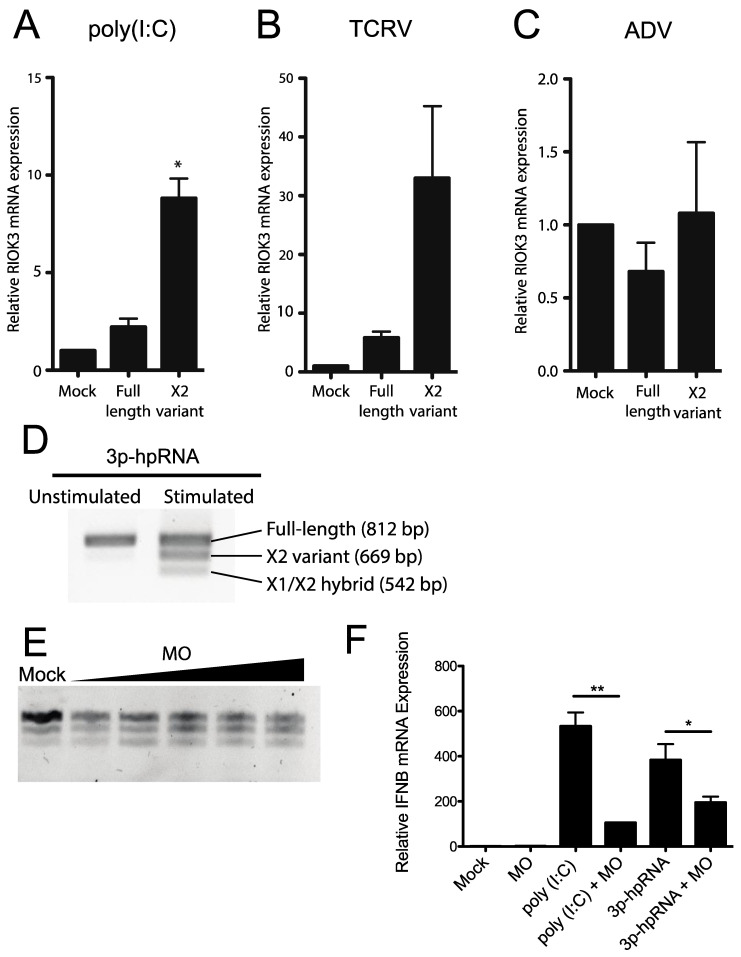
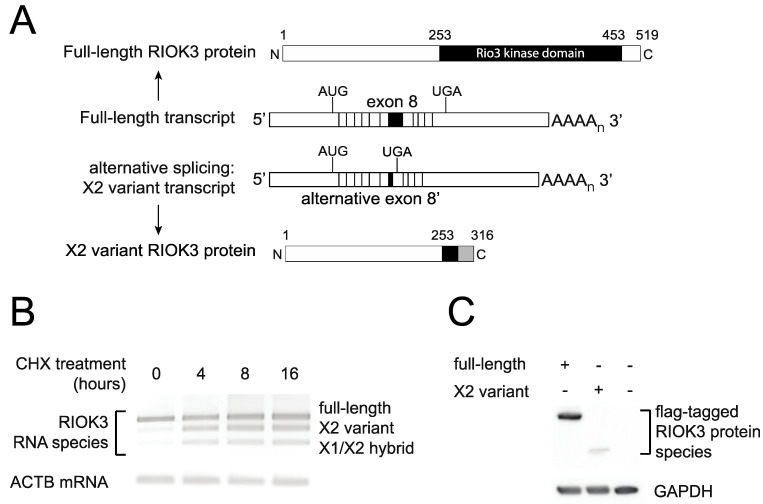
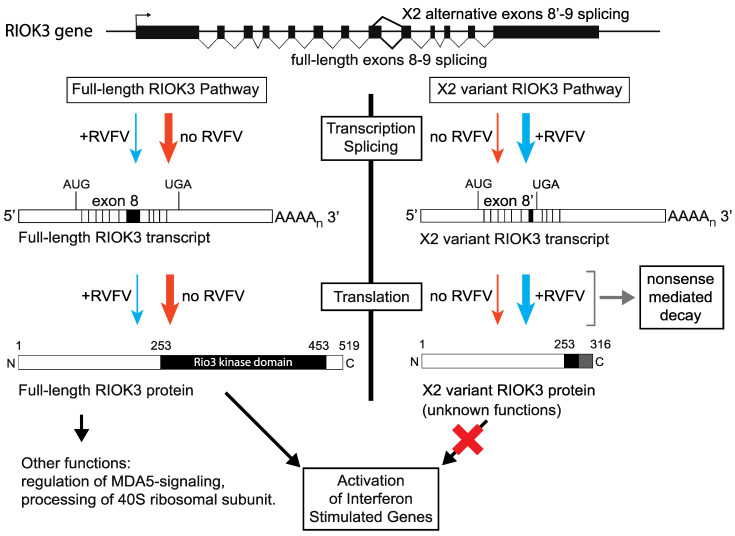
In recent years, transcriptome profiling studies have identified changes in host splicing patterns caused by viral invasion, yet the functional consequences of the vast majority of these splicing events remain uncharacterized. We recently showed that the host splicing landscape changes during Rift Valley fever virus MP-12 strain (RVFV MP-12) infection of mammalian cells. Of particular interest, we observed that the host mRNA for Rio Kinase 3 (RIOK3) was alternatively spliced during infection. This kinase has been shown to be involved in pattern recognition receptor (PRR) signaling mediated by RIG-I like receptors to produce type-I interferon. Here, we characterize RIOK3 as an important component of the interferon signaling pathway during RVFV infection and demonstrate that RIOK3 mRNA expression is skewed shortly after infection to produce alternatively spliced variants that encode premature termination codons. This splicing event plays a critical role in regulation of the antiviral response. Interestingly, infection with other RNA viruses and transfection with nucleic acid-based RIG-I agonists also stimulated RIOK3 alternative splicing. Finally, we show that specifically stimulating alternative splicing of the RIOK3 transcript using a morpholino oligonucleotide reduced interferon expression. Collectively, these results indicate that RIOK3 is an important component of the mammalian interferon signaling cascade and its splicing is a potent regulatory mechanism capable of fine-tuning the host interferon response.
Keywords: MP-12; RIOK3; Rift Valley fever virus; alternative splicing; innate immune response; morpholino oligonucleotide.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Adams M.J., Lefkowitz E.J., King A.M.Q., Harrach B., Harrison R.L., Knowles N.J., Kropinski A.M., Krupovic M., Kuhn J.H., Mushegian A.R., et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2017;162:2505–2538. doi: 10.1007/s00705-017-3358-5. - DOI - PubMed
-
- Coetzer J.A.W., Tustin R.C. Infectious Diseases of Livestock. 2nd ed. Oxford University Press; Cape Town, South Africa: 2004.
-
- Baudin M., Jumaa A.M., Jomma H.J.E., Karsany M.S., Bucht G., Näslund J., Ahlm C., Evander M., Mohamed N. Association of Rift Valley fever virus infection with miscarriage in Sudanese women: A cross-sectional study. Lancet Glob. Health. 2016;4:e864–e871. doi: 10.1016/S2214-109X(16)30176-0. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
