

This is a preprint.
A single-cell and spatial atlas of autopsy tissues reveals pathology and cellular targets of SARS-CoV-2
- PMID: 33655247
- PMCID: PMC7924267
- DOI: 10.1101/2021.02.25.430130
A single-cell and spatial atlas of autopsy tissues reveals pathology and cellular targets of SARS-CoV-2
Abstract
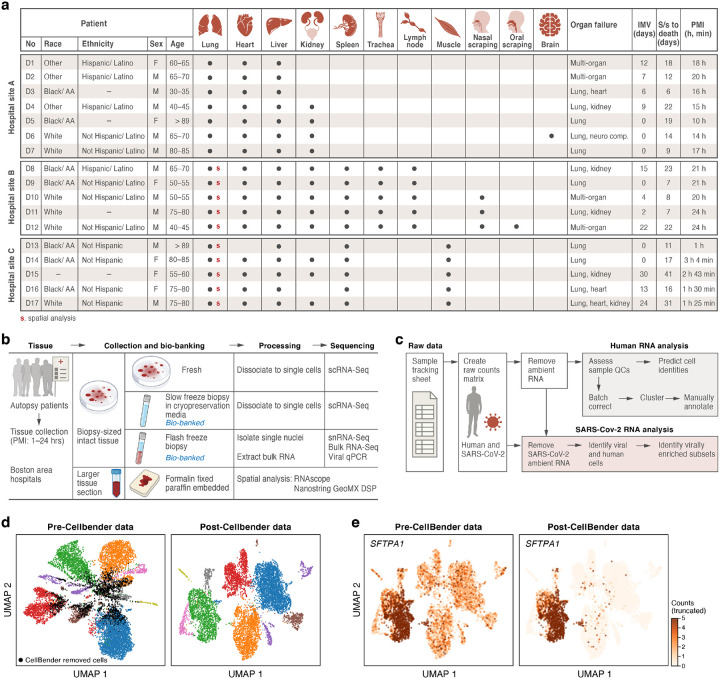
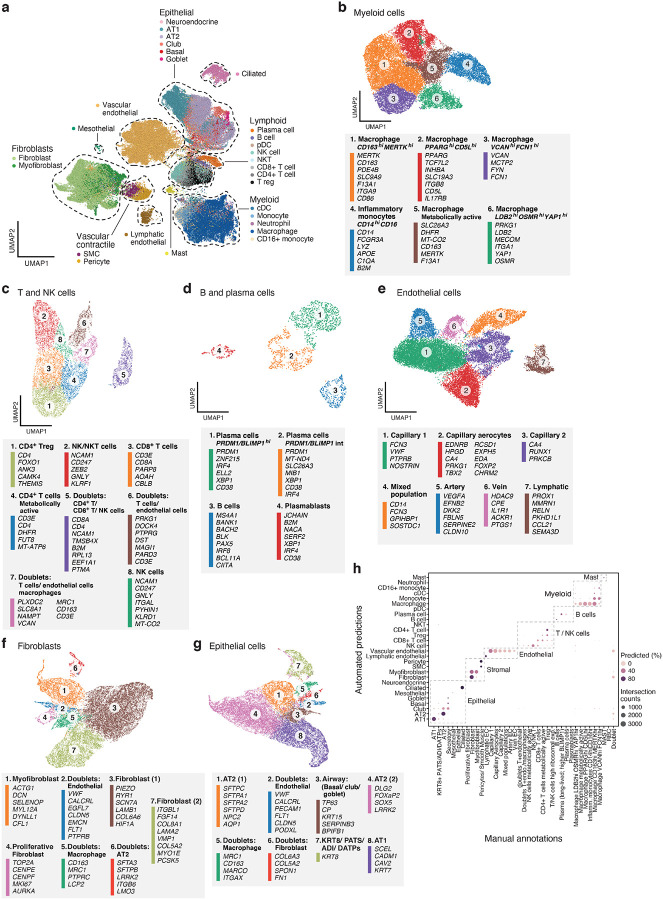
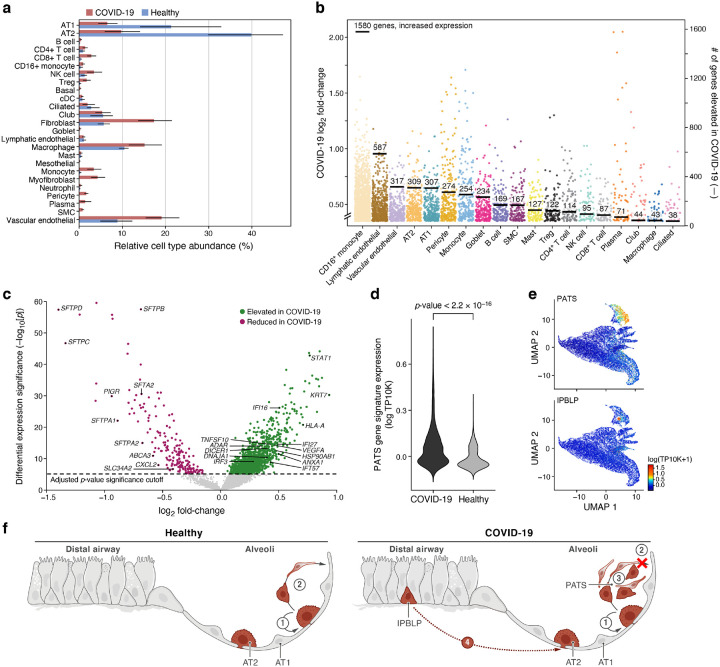
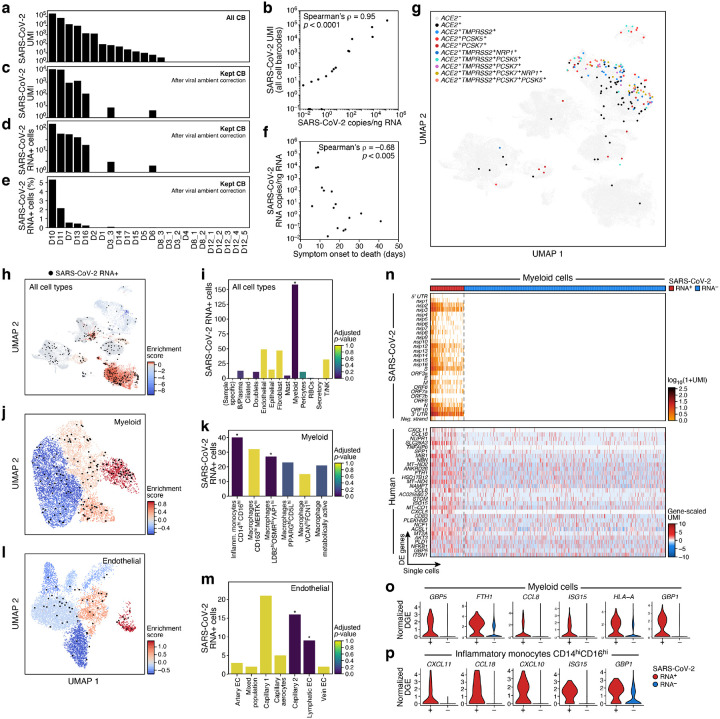
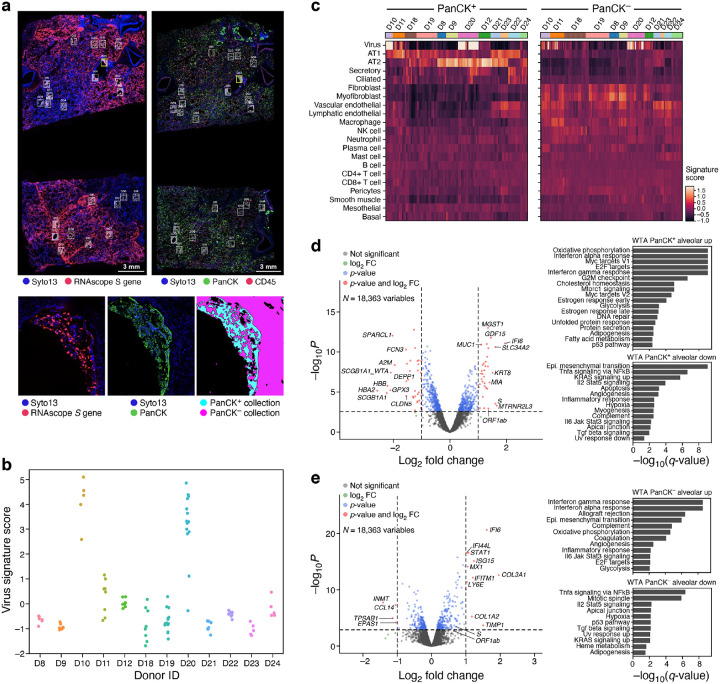
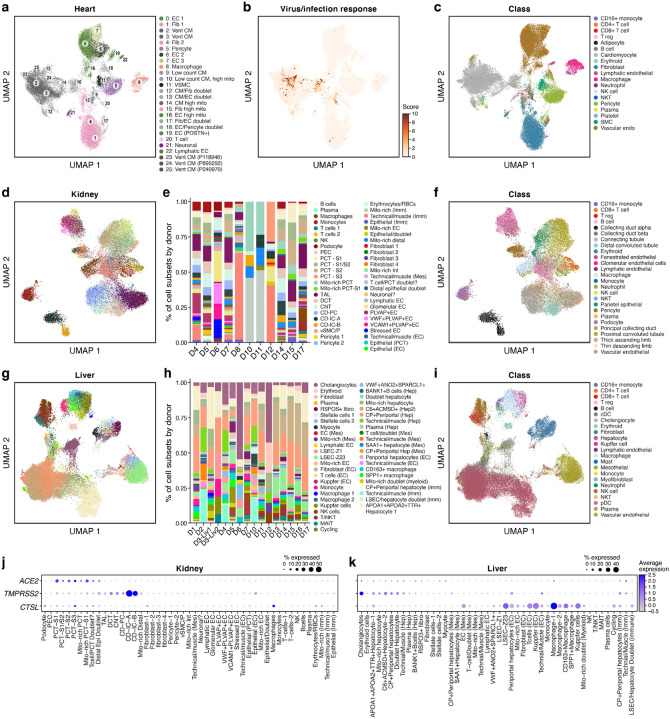
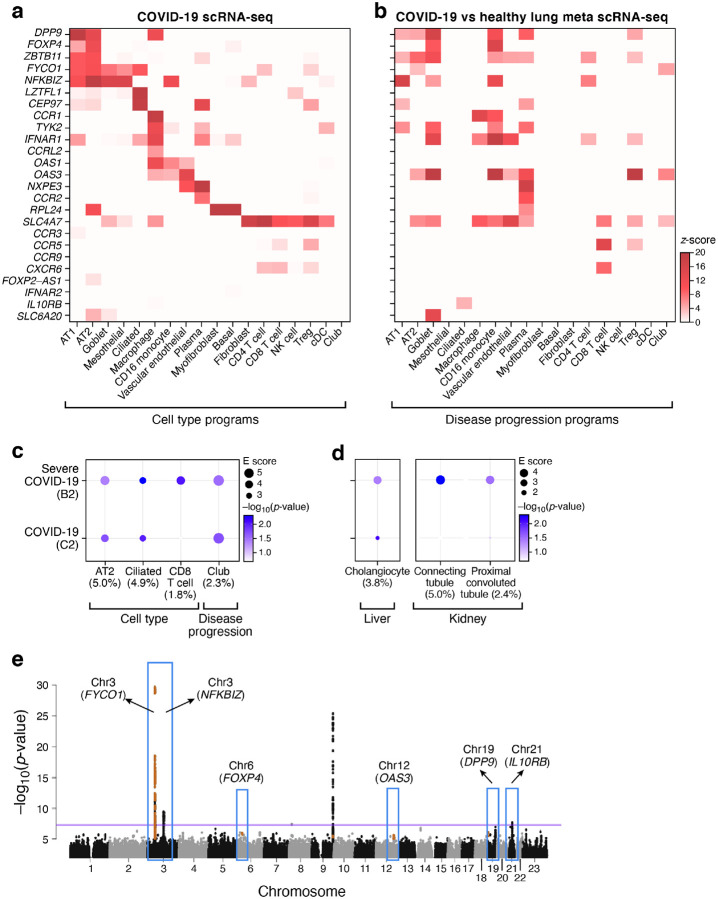
The SARS-CoV-2 pandemic has caused over 1 million deaths globally, mostly due to acute lung injury and acute respiratory distress syndrome, or direct complications resulting in multiple-organ failures. Little is known about the host tissue immune and cellular responses associated with COVID-19 infection, symptoms, and lethality. To address this, we collected tissues from 11 organs during the clinical autopsy of 17 individuals who succumbed to COVID-19, resulting in a tissue bank of approximately 420 specimens. We generated comprehensive cellular maps capturing COVID-19 biology related to patients' demise through single-cell and single-nucleus RNA-Seq of lung, kidney, liver and heart tissues, and further contextualized our findings through spatial RNA profiling of distinct lung regions. We developed a computational framework that incorporates removal of ambient RNA and automated cell type annotation to facilitate comparison with other healthy and diseased tissue atlases. In the lung, we uncovered significantly altered transcriptional programs within the epithelial, immune, and stromal compartments and cell intrinsic changes in multiple cell types relative to lung tissue from healthy controls. We observed evidence of: alveolar type 2 (AT2) differentiation replacing depleted alveolar type 1 (AT1) lung epithelial cells, as previously seen in fibrosis; a concomitant increase in myofibroblasts reflective of defective tissue repair; and, putative TP63+ intrapulmonary basal-like progenitor (IPBLP) cells, similar to cells identified in H1N1 influenza, that may serve as an emergency cellular reserve for severely damaged alveoli. Together, these findings suggest the activation and failure of multiple avenues for regeneration of the epithelium in these terminal lungs. SARS-CoV-2 RNA reads were enriched in lung mononuclear phagocytic cells and endothelial cells, and these cells expressed distinct host response transcriptional programs. We corroborated the compositional and transcriptional changes in lung tissue through spatial analysis of RNA profiles in situ and distinguished unique tissue host responses between regions with and without viral RNA, and in COVID-19 donor tissues relative to healthy lung. Finally, we analyzed genetic regions implicated in COVID-19 GWAS with transcriptomic data to implicate specific cell types and genes associated with disease severity. Overall, our COVID-19 cell atlas is a foundational dataset to better understand the biological impact of SARS-CoV-2 infection across the human body and empowers the identification of new therapeutic interventions and prevention strategies.
Conflict of interest statement
Competing Interests P.D., R.F., E.M.M., M.R., E.H.R., L.P., T.He., J.R., J.B., and S.W. are employees and stockholders at Nanostring Technologies Inc. D.Z., is a former employee and stockholder at NanoString Technologies. N.H., holds equity in BioNTech and Related Sciences. T.H.is an employee and stockholder of Prime Medicine as of Oct. 13, 2020. G.H. is an employee of Genentech as of Nov 16, 2020. R.N. is a founder, shareholder, and member of the board at Rhinostics Inc. A.R. is a co-founder and equity holder of Celsius Therapeutics, an equity holder in Immunitas, and was an SAB member of ThermoFisher Scientific, Syros Pharmaceuticals, Neogene Therapeutics and Asimov until July 31, 2020. From August 1, 2020, A.R. is an employee of Genentech. From October 19, 2020, O.R.-R is an employee of Genentech. P.C.S is a co-founder and shareholder of Sherlock Biosciences, and a Board member and shareholder of Danaher Corporation. A.K.S. reports compensation for consulting and/or SAB membership from Honeycomb Biotechnologies, Cellarity, Repertoire Immune Medicines, Ochre Bio, and Dahlia Biosciences. Z.G.J. reports grant support from Gilead Science, Pfizer, compensation for consulting from Olix Pharmaceuticals. Y.V.P. reports grant support from Enanta Pharmaceuticals, CymaBay Therapeutics, Morphic Therapeutic; consulting and/or SAB in Ambys Medicines, Morphic Therapeutics, Enveda Therapeutics, BridgeBio Pharma, as well as being an Editor – American Journal of Physiology-Gastrointestinal and Liver Physiology. GS reports consultant service in Alnylam Pharmaceuticals, Merck, Generon, Glympse Bio, Inc., Mayday Foundation, Novartis Pharmaceuticals, Quest Diagnostics, Surrozen, Terra Firma, Zomagen Bioscience, Pandion Therapeutics, Inc. Durect Corporation; royalty from UpToDate Inc., and Editor service in Hepatology Communications. P.R.T. receives consulting fees from Cellarity Inc., and Surrozen Inc., for work not related to this manuscript.
Figures
Similar articles
-
COVID-19 tissue atlases reveal SARS-CoV-2 pathology and cellular targets.Nature. 2021 Jul;595(7865):107-113. doi: 10.1038/s41586-021-03570-8. Epub 2021 Apr 29. Nature. 2021. PMID: 33915569 Free PMC article.
-
Molecular and immune signatures, and pathological trajectories of fatal COVID-19 lungs defined by in situ spatial single-cell transcriptome analysis.J Med Virol. 2023 Aug;95(8):e29009. doi: 10.1002/jmv.29009. J Med Virol. 2023. PMID: 37563850 Free PMC article.
-
Subcellular Detection of SARS-CoV-2 RNA in Human Tissue Reveals Distinct Localization in Alveolar Type 2 Pneumocytes and Alveolar Macrophages.mBio. 2021 Feb 22;13(1):e0375121. doi: 10.1128/mbio.03751-21. Epub 2022 Feb 8. mBio. 2021. PMID: 35130722 Free PMC article.
-
Insights into COVID-19 Vaccine Development Based on Immunogenic Structural Proteins of SARS-CoV-2, Host Immune Responses, and Herd Immunity.Cells. 2021 Oct 29;10(11):2949. doi: 10.3390/cells10112949. Cells. 2021. PMID: 34831172 Free PMC article. Review.
-
Function of epithelial stem cell in the repair of alveolar injury.Stem Cell Res Ther. 2022 Apr 27;13(1):170. doi: 10.1186/s13287-022-02847-7. Stem Cell Res Ther. 2022. PMID: 35477551 Free PMC article. Review.
Cited by
-
PlaqView 2.0: A comprehensive web portal for cardiovascular single-cell genomics.Front Cardiovasc Med. 2022 Aug 8;9:969421. doi: 10.3389/fcvm.2022.969421. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 36003902 Free PMC article.
-
miQC: An adaptive probabilistic framework for quality control of single-cell RNA-sequencing data.PLoS Comput Biol. 2021 Aug 24;17(8):e1009290. doi: 10.1371/journal.pcbi.1009290. eCollection 2021 Aug. PLoS Comput Biol. 2021. PMID: 34428202 Free PMC article.
-
Targeting the transmembrane cytokine co-receptor neuropilin-1 in distal tubules improves renal injury and fibrosis.Nat Commun. 2024 Jul 8;15(1):5731. doi: 10.1038/s41467-024-50121-6. Nat Commun. 2024. PMID: 38977708 Free PMC article.
-
GPS tracking analyses reveal finely-tuned shorebird space use and movement patterns throughout the non-breeding season in high-latitude austral intertidal areas.Mov Ecol. 2023 Sep 1;11(1):55. doi: 10.1186/s40462-023-00411-3. Mov Ecol. 2023. PMID: 37658459 Free PMC article.
-
Endothelial cell plasticity at the single-cell level.Angiogenesis. 2021 May;24(2):311-326. doi: 10.1007/s10456-021-09797-3. Epub 2021 Jun 1. Angiogenesis. 2021. PMID: 34061284 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous