

RhoA/Rock activation represents a new mechanism for inactivating Wnt/β-catenin signaling in the aging-associated bone loss
- PMID: 33655459
- PMCID: PMC7925793
- DOI: 10.1186/s13619-020-00071-3
RhoA/Rock activation represents a new mechanism for inactivating Wnt/β-catenin signaling in the aging-associated bone loss
Erratum in
-
Correction: RhoA/Rock activation represents a new mechanism for inactivating Wnt/β-catenin signaling in the aging-associated bone loss.Cell Regen. 2024 Feb 10;13(1):3. doi: 10.1186/s13619-023-00185-4. Cell Regen. 2024. PMID: 38336932 Free PMC article. No abstract available.
Abstract
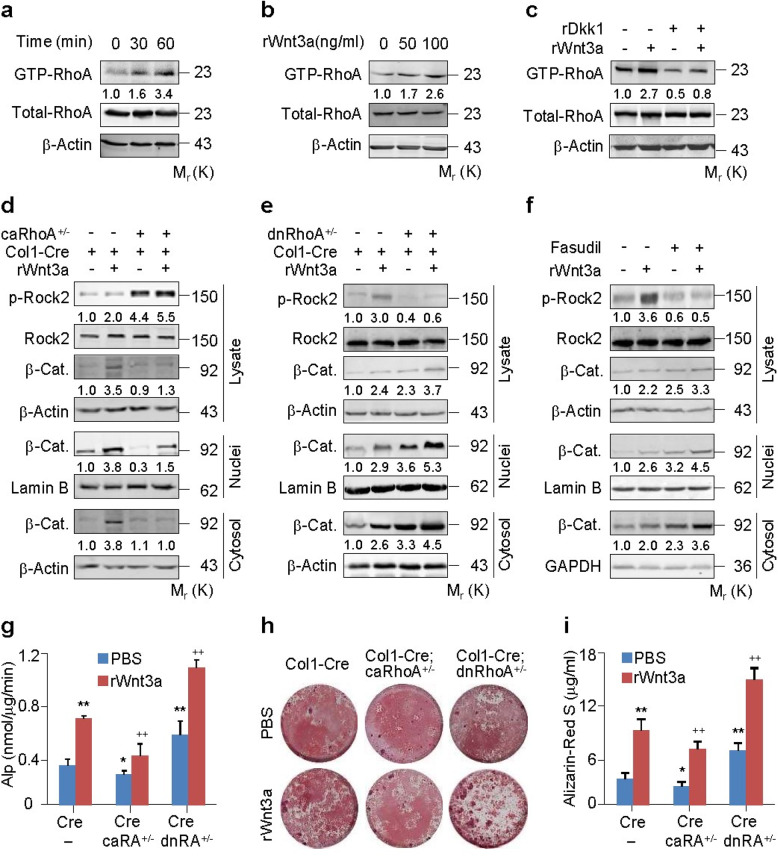
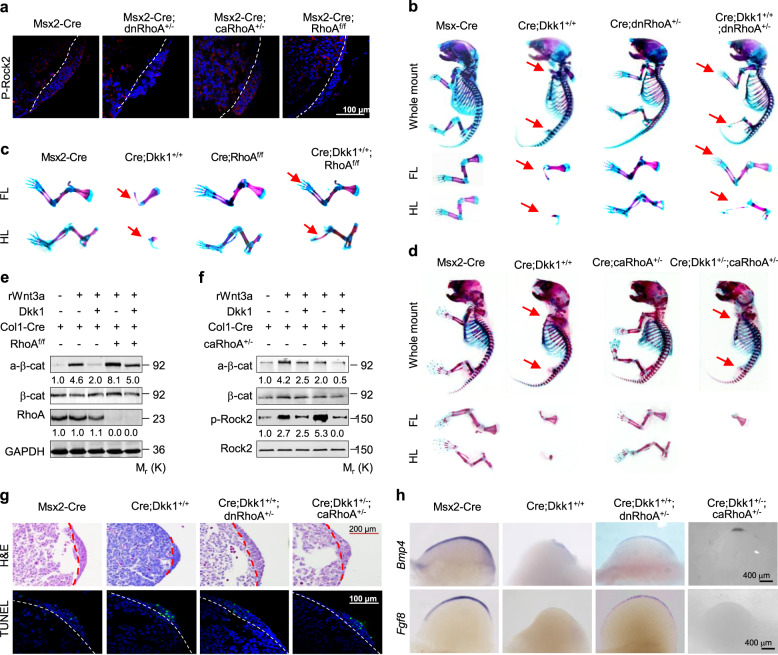
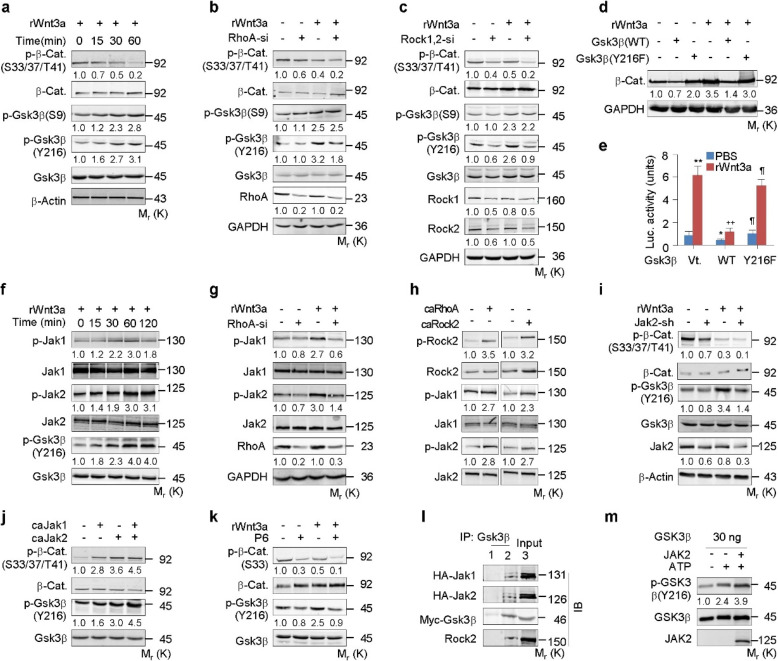
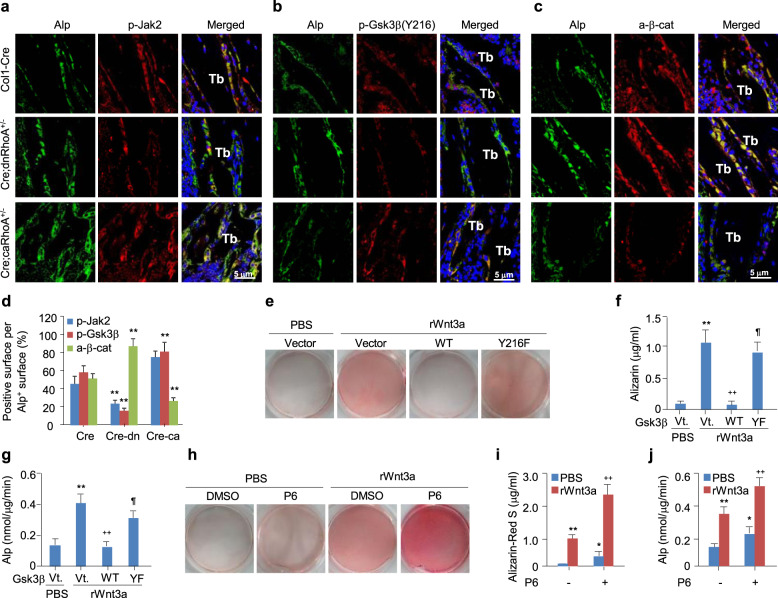
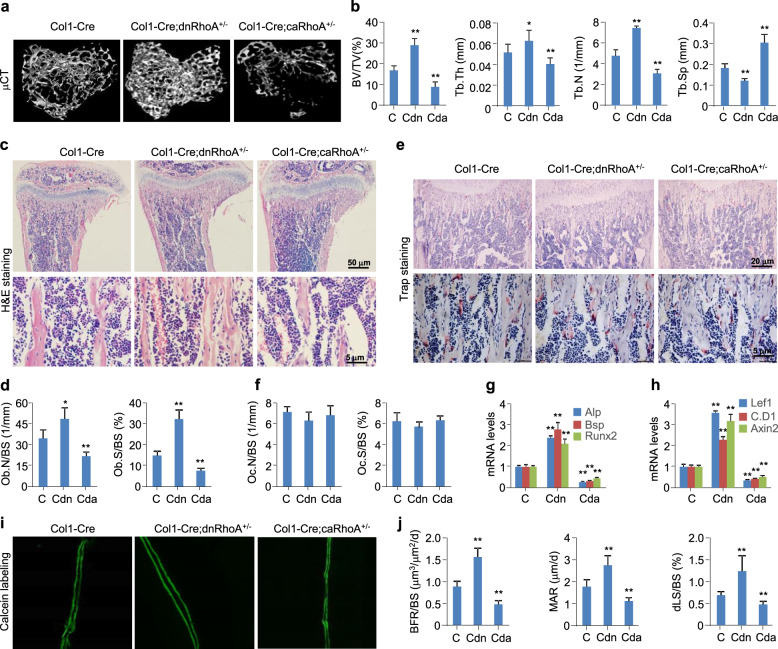
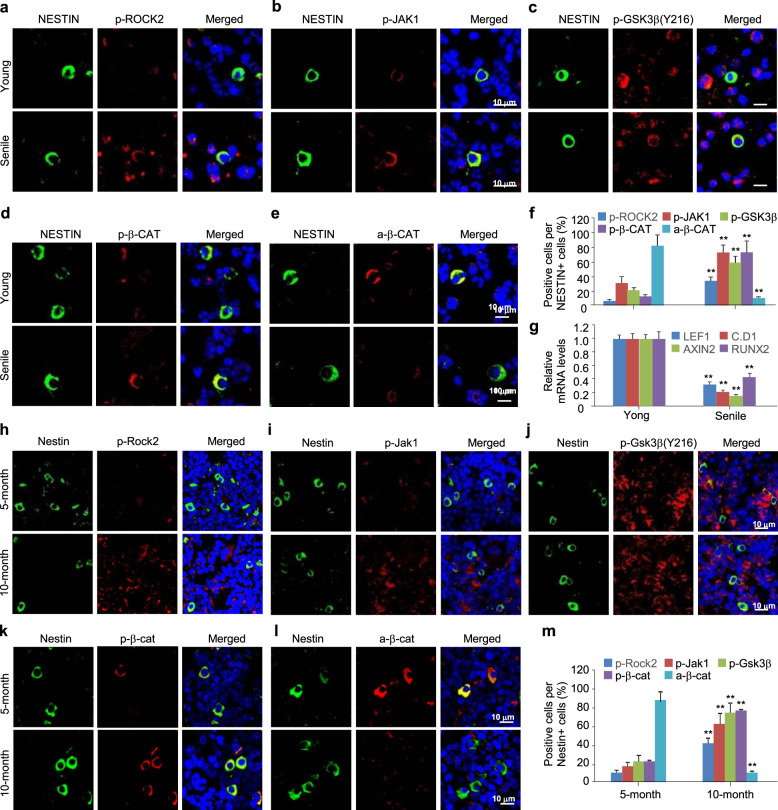
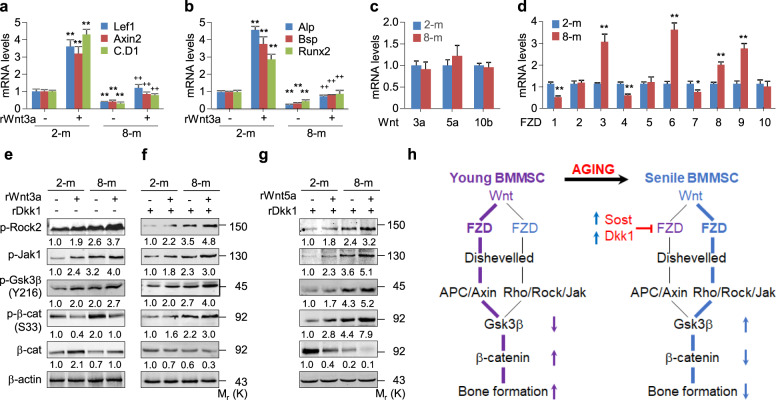
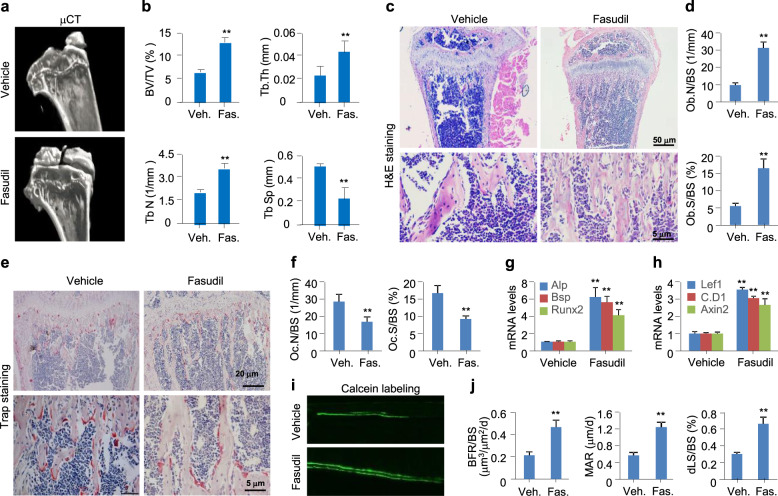
The Wnt/β-catenin signaling pathway appears to be particularly important for bone homeostasis, whereas nuclear accumulation of β-catenin requires the activation of Rac1, a member of the Rho small GTPase family. The aim of the present study was to investigate the role of RhoA/Rho kinase (Rock)-mediated Wnt/β-catenin signaling in the regulation of aging-associated bone loss. We find that Lrp5/6-dependent and Lrp5/6-independent RhoA/Rock activation by Wnt3a activates Jak1/2 to directly phosphorylate Gsk3β at Tyr216, resulting in Gsk3β activation and subsequent β-catenin destabilization. In line with these molecular events, RhoA loss- or gain-of-function in mouse embryonic limb bud ectoderms interacts genetically with Dkk1 gain-of-function to rescue the severe limb truncation phenotypes or to phenocopy the deletion of β-catenin, respectively. Likewise, RhoA loss-of-function in pre-osteoblasts robustly increases bone formation while gain-of-function decreases it. Importantly, high RhoA/Rock activity closely correlates with Jak and Gsk3β activities but inversely correlates with β-catenin signaling activity in bone marrow mesenchymal stromal cells from elderly male humans and mice, whereas systemic inhibition of Rock therefore activates the β-catenin signaling to antagonize aging-associated bone loss. Taken together, these results identify RhoA/Rock-dependent Gsk3β activation and subsequent β-catenin destabilization as a hitherto uncharacterized mechanism controlling limb outgrowth and bone homeostasis.
Keywords: Bone; Limb bud; RhoA; Rock; Wnt; β-Catenin.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous