

Astrocytes are HIV reservoirs in the brain: A cell type with poor HIV infectivity and replication but efficient cell-to-cell viral transfer
- PMID: 33655498
- PMCID: PMC11102126
- DOI: 10.1111/jnc.15336
Astrocytes are HIV reservoirs in the brain: A cell type with poor HIV infectivity and replication but efficient cell-to-cell viral transfer
Abstract
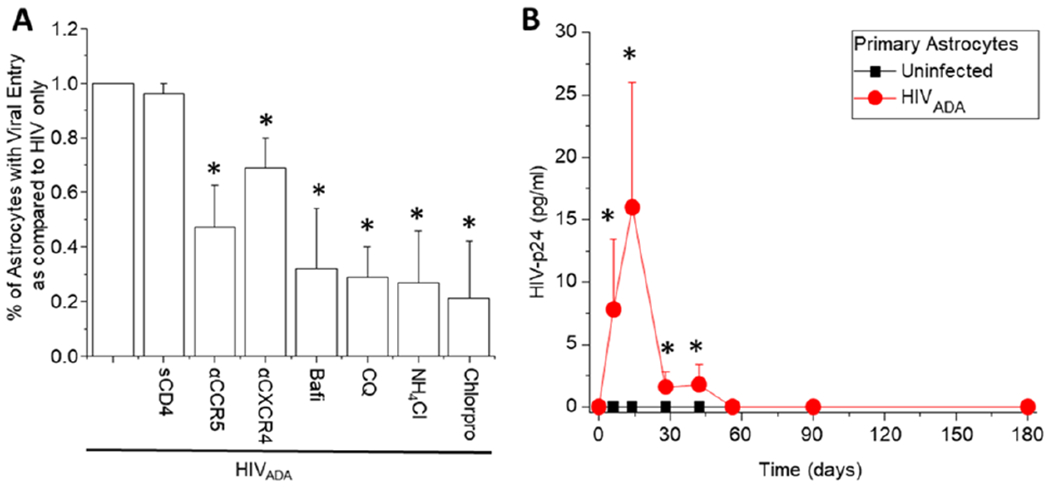
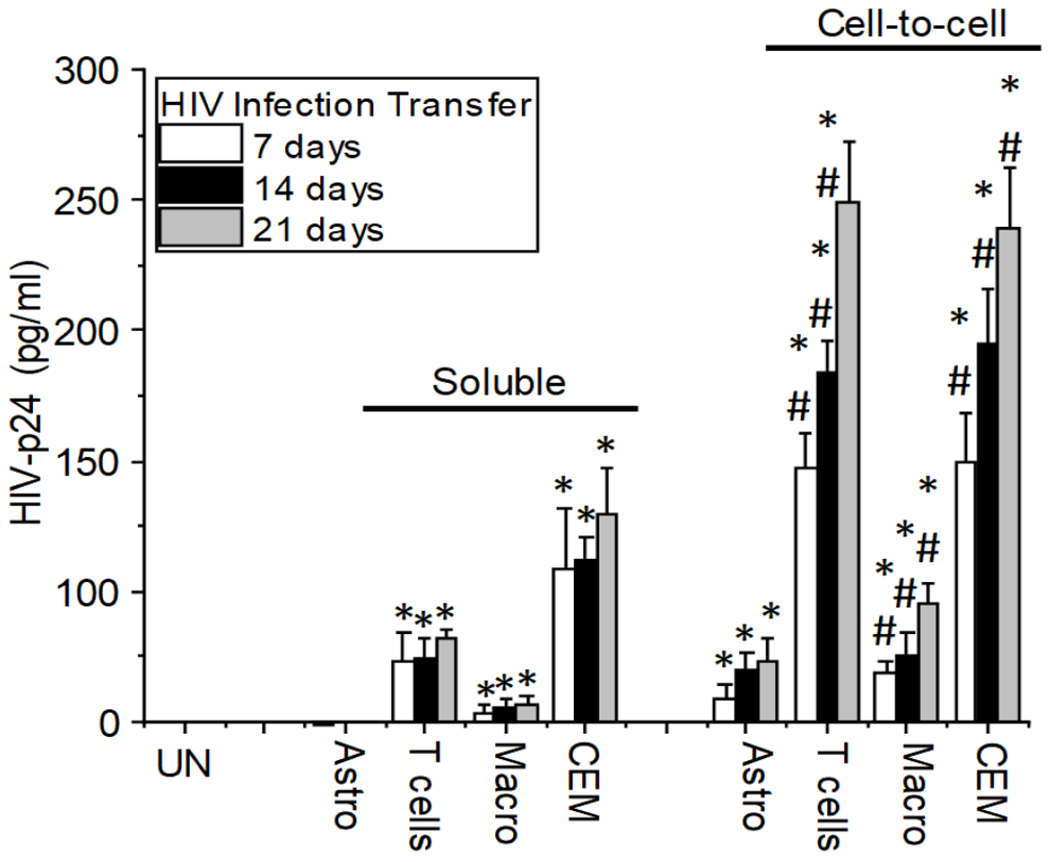
The major barrier to eradicating Human immunodeficiency virus-1 (HIV) infection is the generation of tissue-associated quiescent long-lasting viral reservoirs refractory to therapy. Upon interruption of anti-retroviral therapy (ART), HIV replication can be reactivated. Within the brain, microglia/macrophages and a small population of astrocytes are infected with HIV. However, the role of astrocytes as a potential viral reservoir is becoming more recognized because of the improved detection and quantification of HIV viral reservoirs. In this report, we examined the infectivity of human primary astrocytes in vivo and in vitro, and their capacity to maintain HIV infection, become latently infected, be reactivated, and transfer new HIV virions into neighboring cells. Analysis of human brain tissue sections obtained from HIV-infected individuals under effective and prolonged ART indicates that a small population of astrocytes has integrated HIV-DNA. In vitro experiments using HIV-infected human primary astrocyte cultures confirmed a low percentage of astrocytes had integrated HIV-DNA, with poor to undetectable replication. Even in the absence of ART, long-term culture results in latency that could be transiently reactivated with histone deacetylase inhibitor, tumor necrosis factor-alpha (TNF-α), or methamphetamine. Reactivation resulted in poor viral production but efficient cell-to-cell viral transfer into cells that support high viral replication. Together, our data provide a new understanding of astrocytes' role as viral reservoirs within the central nervous system (CNS).
Keywords: NeuroHIV; anti-retroviral; astrocytes; cure; latency; reactivation; reservoirs.
© 2021 International Society for Neurochemistry.
Figures


Similar articles
-
Brain Macrophages in Simian Immunodeficiency Virus-Infected, Antiretroviral-Suppressed Macaques: a Functional Latent Reservoir.mBio. 2017 Aug 15;8(4):e01186-17. doi: 10.1128/mBio.01186-17. mBio. 2017. PMID: 28811349 Free PMC article.
-
Epigenetic Suppression of HIV in Myeloid Cells by the BRD4-Selective Small Molecule Modulator ZL0580.J Virol. 2020 May 18;94(11):e01880-19. doi: 10.1128/JVI.01880-19. Print 2020 May 18. J Virol. 2020. PMID: 32188727 Free PMC article.
-
Quiescence Promotes Latent HIV Infection and Resistance to Reactivation from Latency with Histone Deacetylase Inhibitors.J Virol. 2017 Nov 30;91(24):e01080-17. doi: 10.1128/JVI.01080-17. Print 2017 Dec 15. J Virol. 2017. PMID: 29021396 Free PMC article.
-
Brain macrophages harbor latent, infectious simian immunodeficiency virus.AIDS. 2019 Dec 1;33 Suppl 2(Suppl 2):S181-S188. doi: 10.1097/QAD.0000000000002269. AIDS. 2019. PMID: 31789817 Free PMC article. Review.
-
Reservoirs for HIV-1: mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy.Annu Rev Immunol. 2000;18:665-708. doi: 10.1146/annurev.immunol.18.1.665. Annu Rev Immunol. 2000. PMID: 10837072 Review.
Cited by
-
Neurological, Behavioral, and Pathophysiological Characterization of the Co-Occurrence of Substance Use and HIV: A Narrative Review.Brain Sci. 2023 Oct 19;13(10):1480. doi: 10.3390/brainsci13101480. Brain Sci. 2023. PMID: 37891847 Free PMC article. Review.
-
The role of tunneling nanotubes during early stages of HIV infection and reactivation: implications in HIV cure.NeuroImmune Pharm Ther. 2023 Jan 4;2(2):169-186. doi: 10.1515/nipt-2022-0015. eCollection 2023 Jun. NeuroImmune Pharm Ther. 2023. PMID: 37476291 Free PMC article. Review.
-
Tenofovir Activation is Diminished in the Brain and Liver of Creatine Kinase Brain-Type Knockout Mice.bioRxiv [Preprint]. 2023 Sep 26:2023.09.25.559370. doi: 10.1101/2023.09.25.559370. bioRxiv. 2023. Update in: ACS Pharmacol Transl Sci. 2024 Jan 03;7(1):222-235. doi: 10.1021/acsptsci.3c00250. PMID: 37808667 Free PMC article. Updated. Preprint.
-
Confound, Cause, or Cure: The Effect of Cannabinoids on HIV-Associated Neurological Sequelae.Viruses. 2021 Jun 26;13(7):1242. doi: 10.3390/v13071242. Viruses. 2021. PMID: 34206839 Free PMC article. Review.
-
Pharmacological advances in HIV treatment: from ART to long-acting injectable therapies.Arch Virol. 2025 Aug 19;170(9):195. doi: 10.1007/s00705-025-06381-8. Arch Virol. 2025. PMID: 40826301 Free PMC article. Review.
References
-
- Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, Jacob Filho W, Lent R and Herculano-Houzel S (2009) Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of comparative neurology, 513, 532–541. - PubMed
-
- Barat C, Proust A, Deshiere A, Leboeuf M, Drouin J and Tremblay MJ (2018) Astrocytes sustain long-term productive HIV-1 infection without establishment of reactivable viral latency. Glia, 66, 1363–1381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
