

GRK2 regulates GLP-1R-mediated early phase insulin secretion in vivo
- PMID: 33658023
- PMCID: PMC7931601
- DOI: 10.1186/s12915-021-00966-w
GRK2 regulates GLP-1R-mediated early phase insulin secretion in vivo
Abstract
Background: Insulin secretion from the pancreatic β-cell is finely modulated by different signals to allow an adequate control of glucose homeostasis. Incretin hormones such as glucagon-like peptide-1 (GLP-1) act as key physiological potentiators of insulin release through binding to the G protein-coupled receptor GLP-1R. Another key regulator of insulin signaling is the Ser/Thr kinase G protein-coupled receptor kinase 2 (GRK2). However, whether GRK2 affects insulin secretion or if GRK2 can control incretin actions in vivo remains to be analyzed.
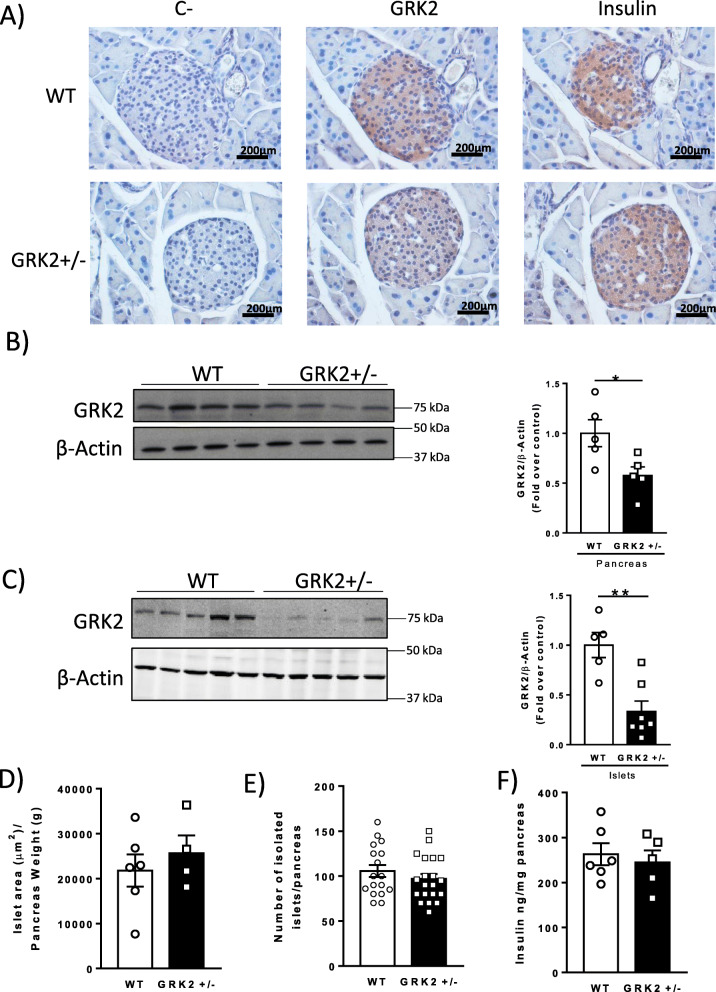
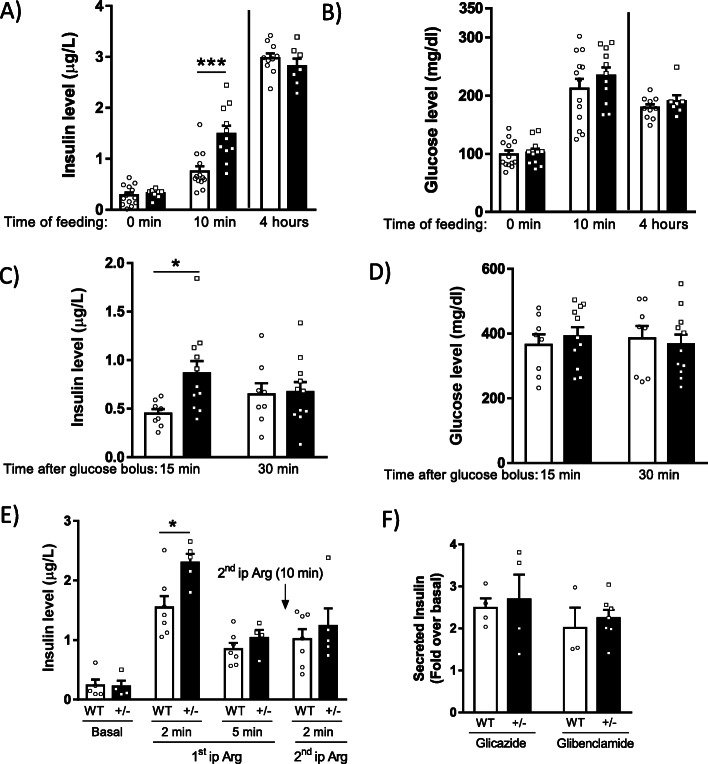
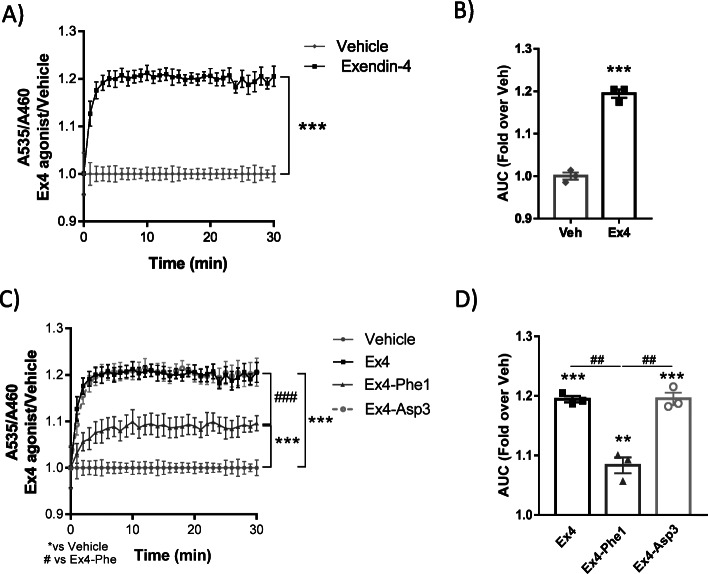
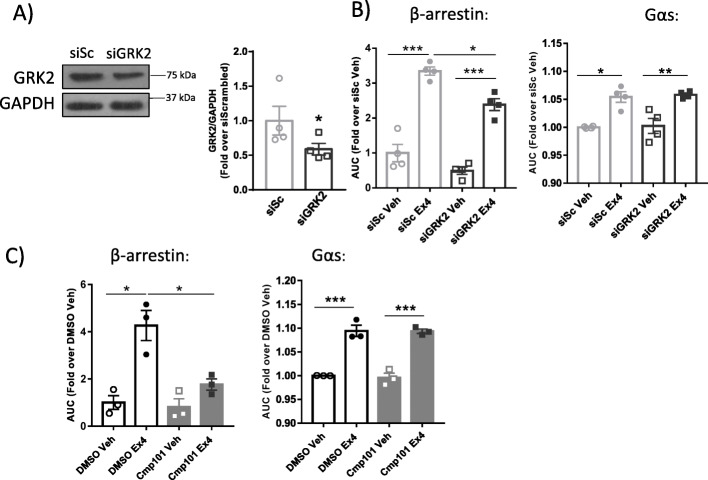
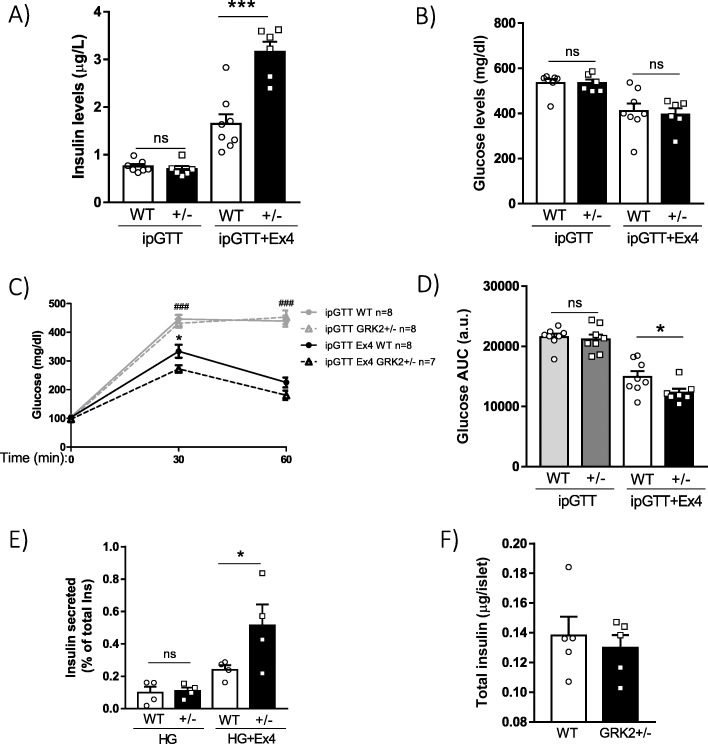
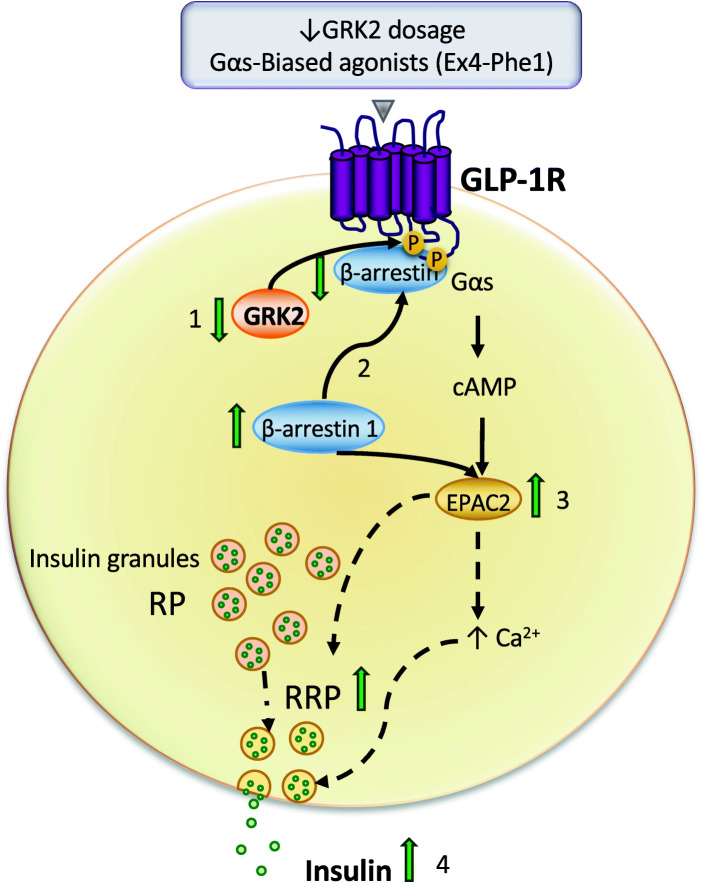
Results: Using GRK2 hemizygous mice, isolated pancreatic islets, and model β-cell lines, we have uncovered a relevant physiological role for GRK2 as a regulator of incretin-mediated insulin secretion in vivo. Feeding, oral glucose gavage, or administration of GLP-1R agonists in animals with reduced GRK2 levels (GRK2+/- mice) resulted in enhanced early phase insulin release without affecting late phase secretion. In contrast, intraperitoneal glucose-induced insulin release was not affected. This effect was recapitulated in isolated islets and correlated with the increased size or priming efficacy of the readily releasable pool (RRP) of insulin granules that was observed in GRK2+/- mice. Using nanoBRET in β-cell lines, we found that stimulation of GLP-1R promoted GRK2 association to this receptor and that GRK2 protein and kinase activity were required for subsequent β-arrestin recruitment.
Conclusions: Overall, our data suggest that GRK2 is an important negative modulator of GLP-1R-mediated insulin secretion and that GRK2-interfering strategies may favor β-cell insulin secretion specifically during the early phase, an effect that may carry interesting therapeutic applications.
Keywords: G protein-coupled receptor kinase 2 (GRK2); Glucagon-like peptide 1 (GLP-1); Granule dynamics; Incretin; Insulin signaling; β-arrestin.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
