

Global profiling of distinct cysteine redox forms reveals wide-ranging redox regulation in C. elegans
- PMID: 33658510
- PMCID: PMC7930113
- DOI: 10.1038/s41467-021-21686-3
Global profiling of distinct cysteine redox forms reveals wide-ranging redox regulation in C. elegans
Abstract
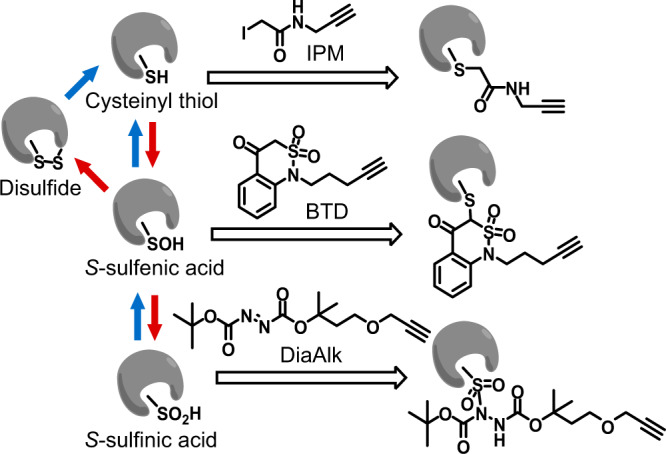
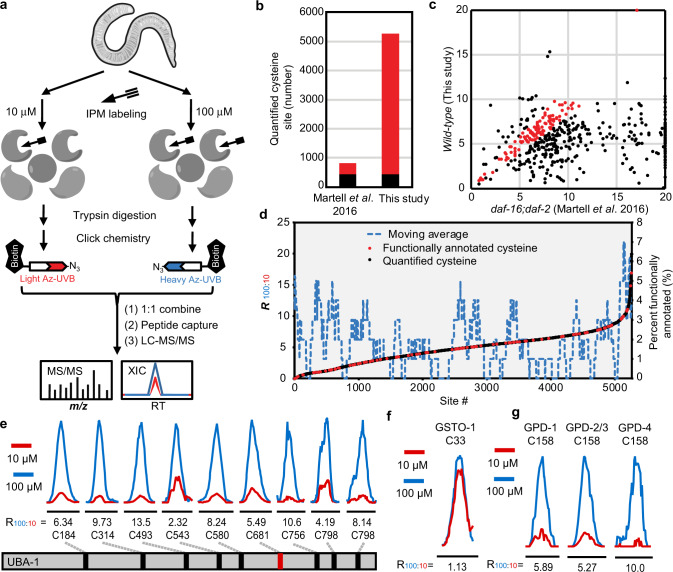
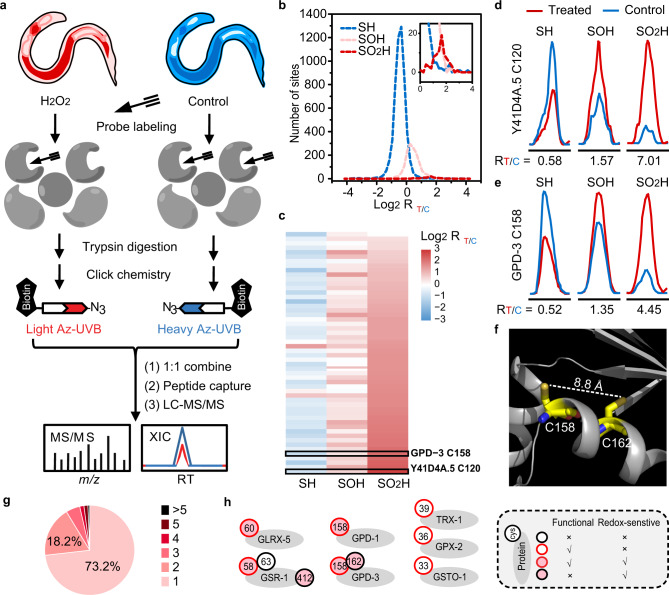
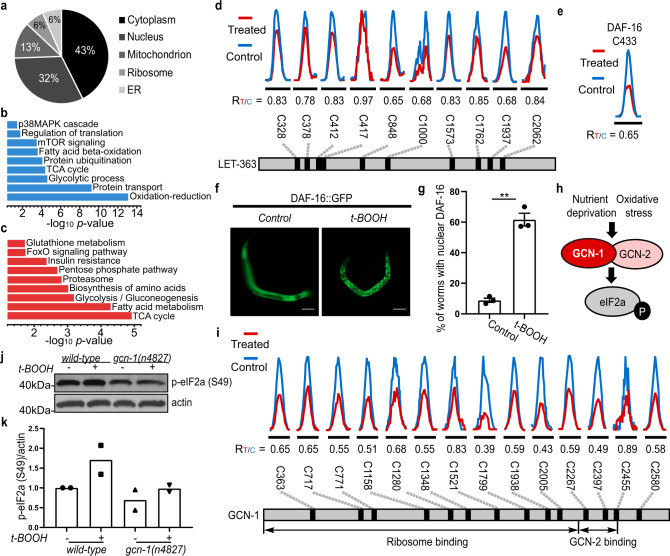
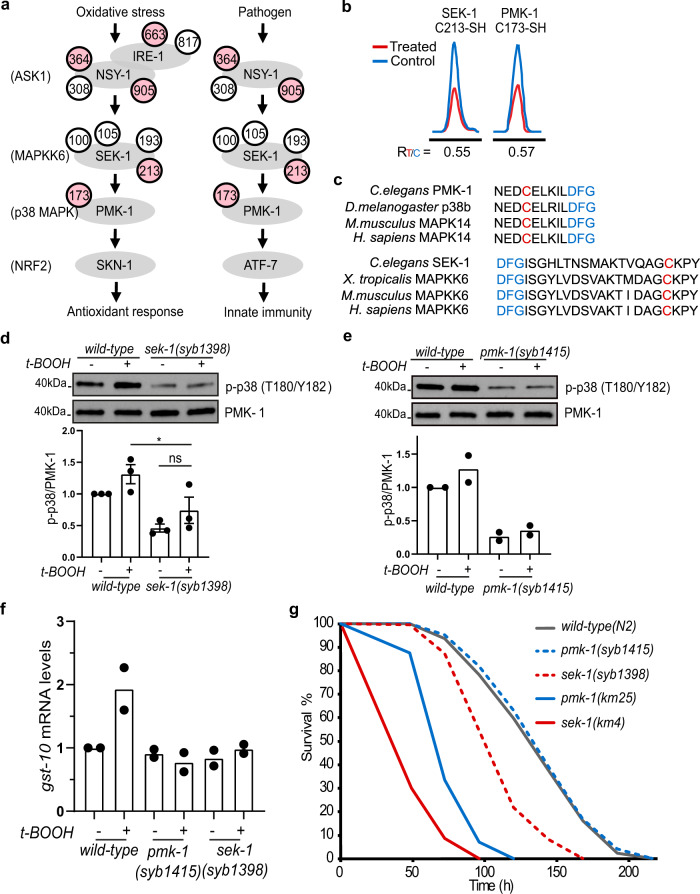
Post-translational changes in the redox state of cysteine residues can rapidly and reversibly alter protein functions, thereby modulating biological processes. The nematode C. elegans is an ideal model organism for studying cysteine-mediated redox signaling at a network level. Here we present a comprehensive, quantitative, and site-specific profile of the intrinsic reactivity of the cysteinome in wild-type C. elegans. We also describe a global characterization of the C. elegans redoxome in which we measured changes in three major cysteine redox forms after H2O2 treatment. Our data revealed redox-sensitive events in translation, growth signaling, and stress response pathways, and identified redox-regulated cysteines that are important for signaling through the p38 MAP kinase (MAPK) pathway. Our in-depth proteomic dataset provides a molecular basis for understanding redox signaling in vivo, and will serve as a valuable and rich resource for the field of redox biology.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Caenorhabditis elegans mounts a p38 MAPK pathway-mediated defence to Cutibacterium acnes infection.Cell Microbiol. 2020 Oct;22(10):e13234. doi: 10.1111/cmi.13234. Epub 2020 Jul 15. Cell Microbiol. 2020. PMID: 32543022
-
The activation of the oxidative stress response transcription factor SKN-1 in Caenorhabditis elegans by mitis group streptococci.PLoS One. 2018 Aug 16;13(8):e0202233. doi: 10.1371/journal.pone.0202233. eCollection 2018. PLoS One. 2018. PMID: 30114261 Free PMC article.
-
Regulation of the Response of Caenorhabditis elegans to Simulated Microgravity by p38 Mitogen-Activated Protein Kinase Signaling.Sci Rep. 2018 Jan 16;8(1):857. doi: 10.1038/s41598-018-19377-z. Sci Rep. 2018. PMID: 29339777 Free PMC article.
-
Factors regulating axon regeneration via JNK MAP kinase in Caenorhabditis elegans.J Biochem. 2020 May 1;167(5):433-439. doi: 10.1093/jb/mvaa020. J Biochem. 2020. PMID: 32091576 Review.
-
Reversible cysteine oxidation in hydrogen peroxide sensing and signal transduction.Biochemistry. 2014 Apr 29;53(16):2560-80. doi: 10.1021/bi401700f. Epub 2014 Apr 16. Biochemistry. 2014. PMID: 24738931 Review.
Cited by
-
Stimulation of RAS-dependent ROS signaling extends longevity by modulating a developmental program of global gene expression.Sci Adv. 2022 Dec 2;8(48):eadc9851. doi: 10.1126/sciadv.adc9851. Epub 2022 Nov 30. Sci Adv. 2022. PMID: 36449615 Free PMC article.
-
Oxidative Stress in Age-Related Neurodegenerative Diseases: An Overview of Recent Tools and Findings.Antioxidants (Basel). 2023 Jan 5;12(1):131. doi: 10.3390/antiox12010131. Antioxidants (Basel). 2023. PMID: 36670993 Free PMC article. Review.
-
Peroxiredoxin 2 is required for the redox mediated adaptation to exercise.Redox Biol. 2023 Apr;60:102631. doi: 10.1016/j.redox.2023.102631. Epub 2023 Feb 9. Redox Biol. 2023. PMID: 36791646 Free PMC article.
-
Redox signaling-mediated tumor extracellular matrix remodeling: pleiotropic regulatory mechanisms.Cell Oncol (Dordr). 2024 Apr;47(2):429-445. doi: 10.1007/s13402-023-00884-9. Epub 2023 Oct 4. Cell Oncol (Dordr). 2024. PMID: 37792154 Review.
-
More questions than answers: insights into potential cysteine-rich receptor-like kinases redox signalling in Arabidopsis.Plant J. 2025 Apr;122(2):e70176. doi: 10.1111/tpj.70176. Plant J. 2025. PMID: 40300149 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
