

Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation
- PMID: 33658521
- PMCID: PMC7930111
- DOI: 10.1038/s41467-021-21718-y
Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation
Abstract
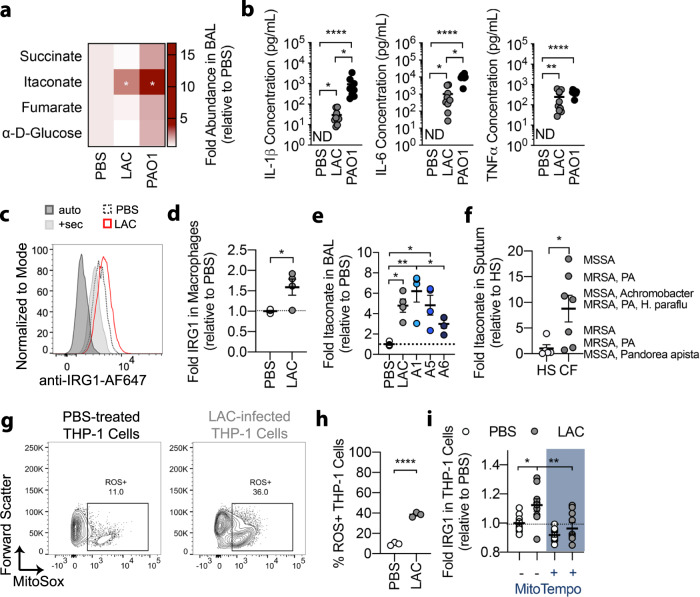
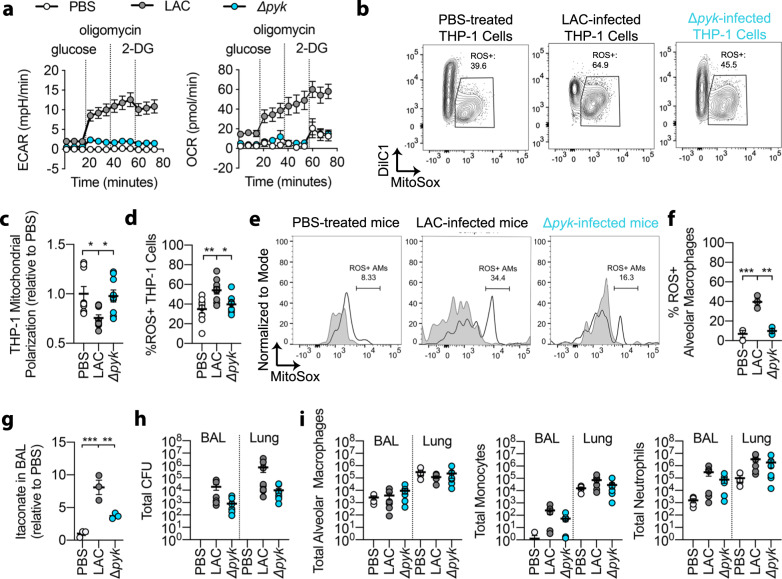
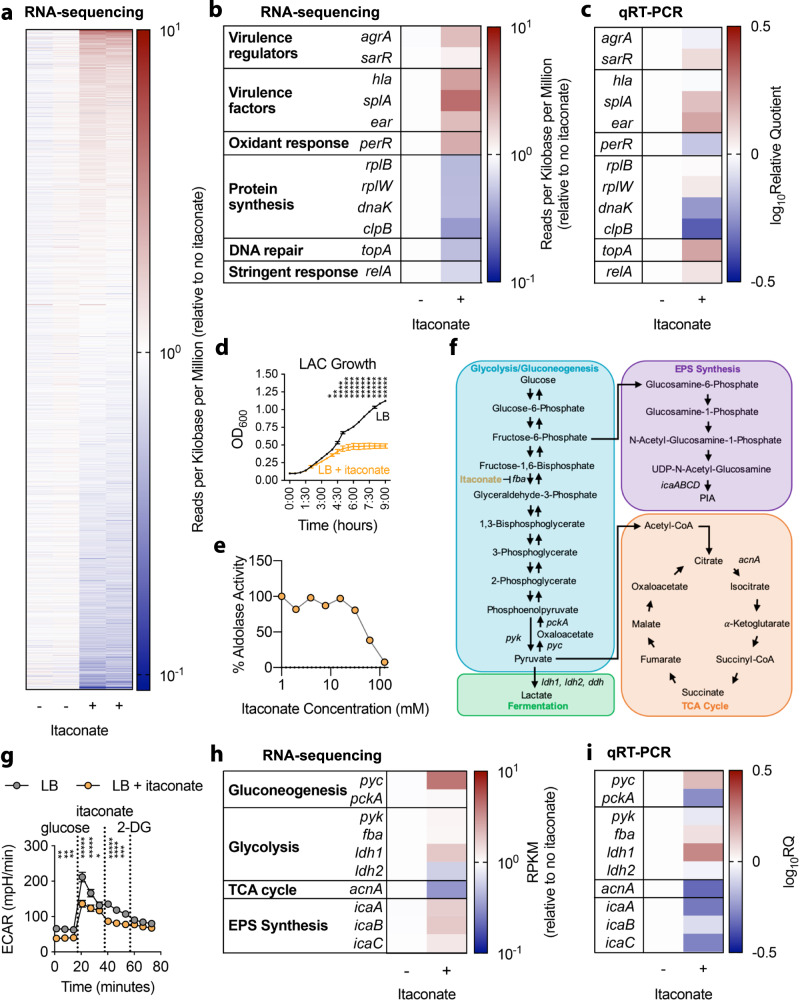
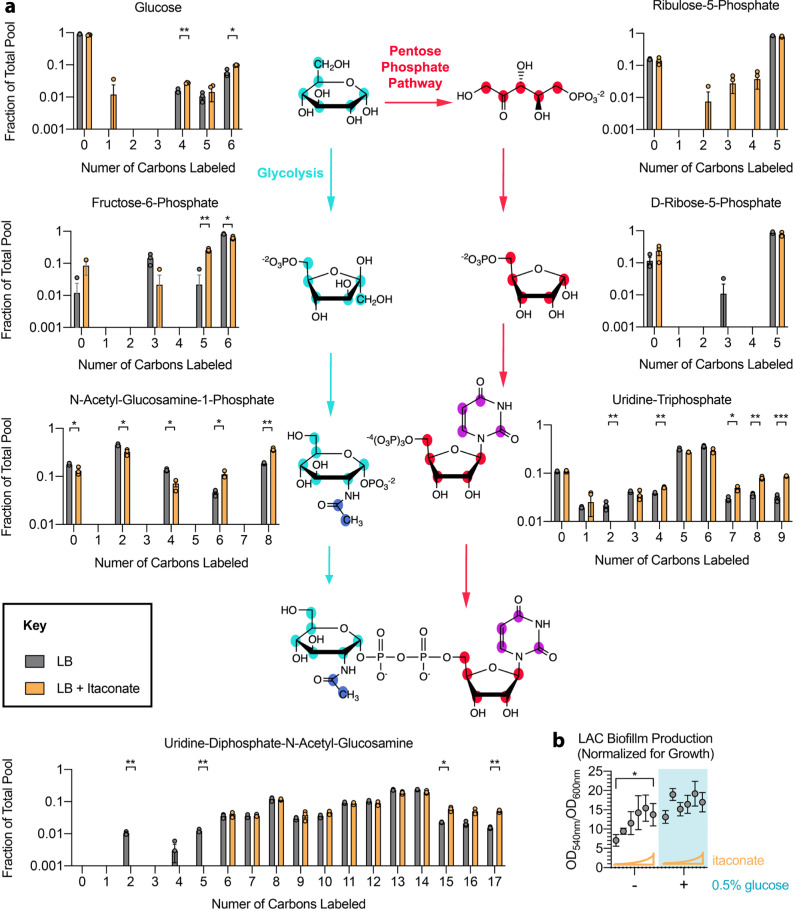
Staphylococcus aureus is a prominent human pathogen that readily adapts to host immune defenses. Here, we show that, in contrast to Gram-negative pathogens, S. aureus induces a distinct airway immunometabolic response dominated by the release of the electrophilic metabolite, itaconate. The itaconate synthetic enzyme, IRG1, is activated by host mitochondrial stress, which is induced by staphylococcal glycolysis. Itaconate inhibits S. aureus glycolysis and selects for strains that re-direct carbon flux to fuel extracellular polysaccharide (EPS) synthesis and biofilm formation. Itaconate-adapted strains, as illustrated by S. aureus isolates from chronic airway infection, exhibit decreased glycolytic activity, high EPS production, and proficient biofilm formation even before itaconate stimulation. S. aureus thus adapts to the itaconate-dominated immunometabolic response by producing biofilms, which are associated with chronic infection of the human airway.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Host-bacteria metabolic crosstalk drives S. aureus biofilm.Microb Cell. 2021 Apr 19;8(5):106-107. doi: 10.15698/mic2021.05.749. Microb Cell. 2021. PMID: 33981762 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
