

Hunger enhances food-odour attraction through a neuropeptide Y spotlight
- PMID: 33658716
- PMCID: PMC8035273
- DOI: 10.1038/s41586-021-03299-4
Hunger enhances food-odour attraction through a neuropeptide Y spotlight
Abstract
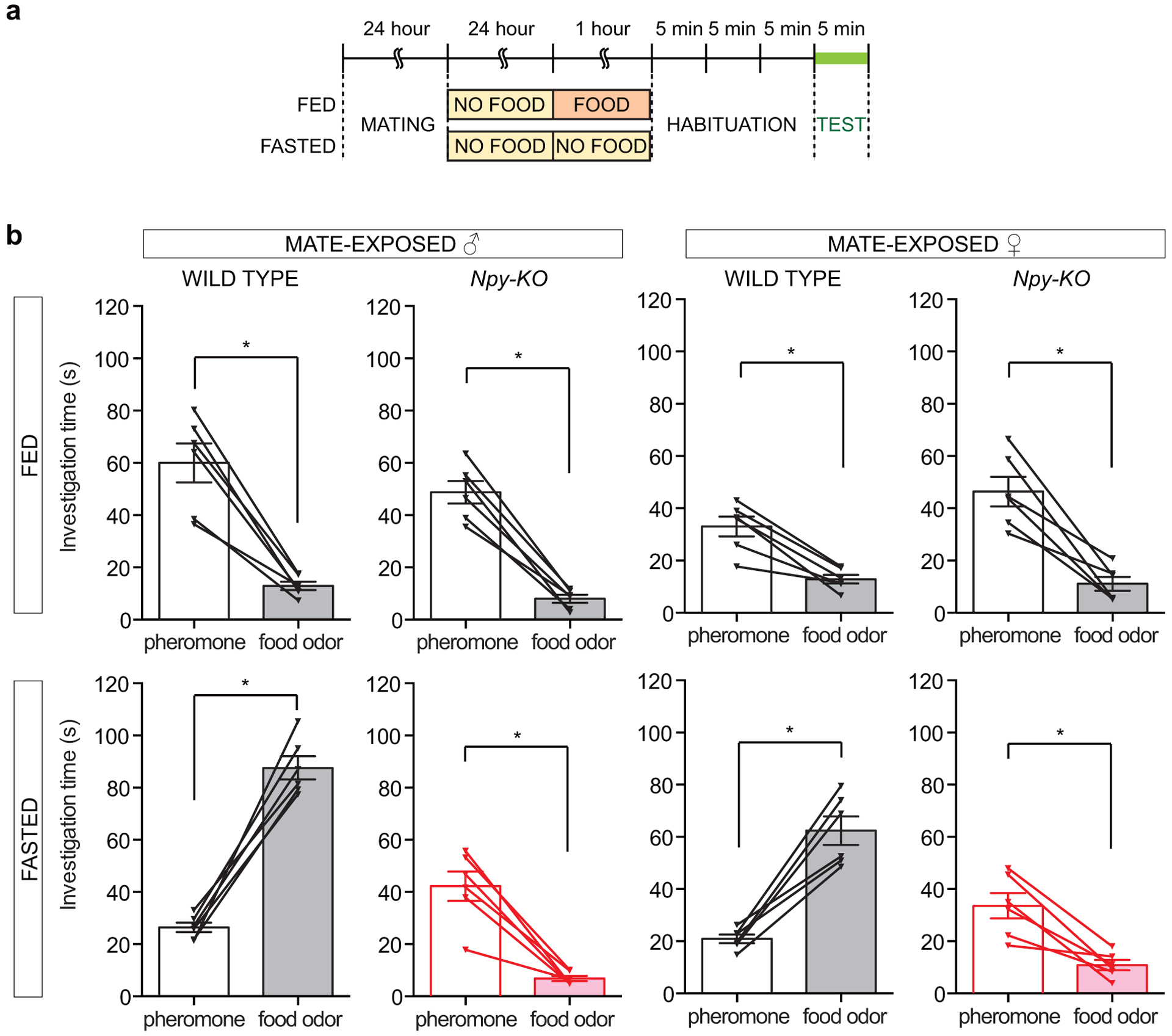
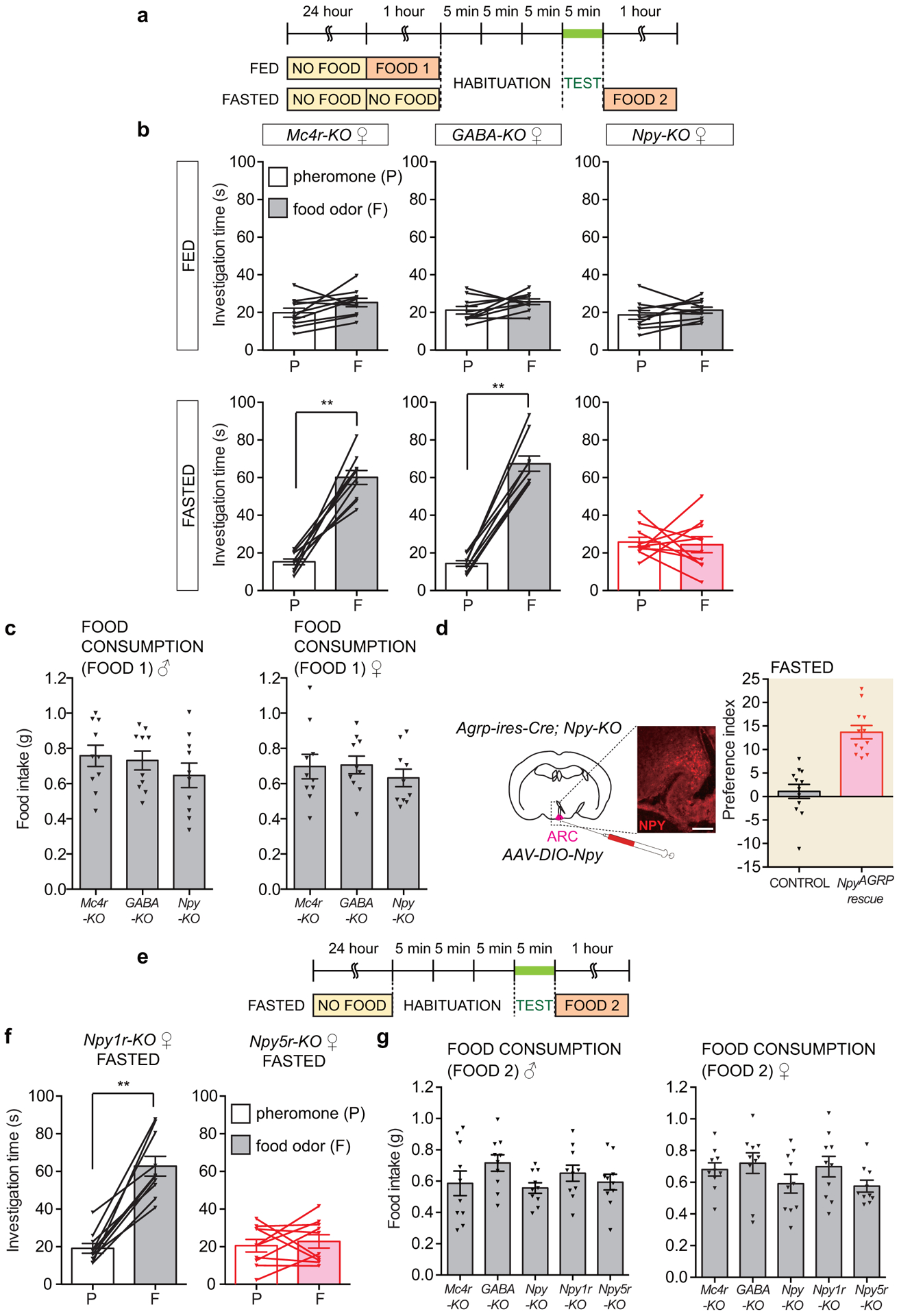
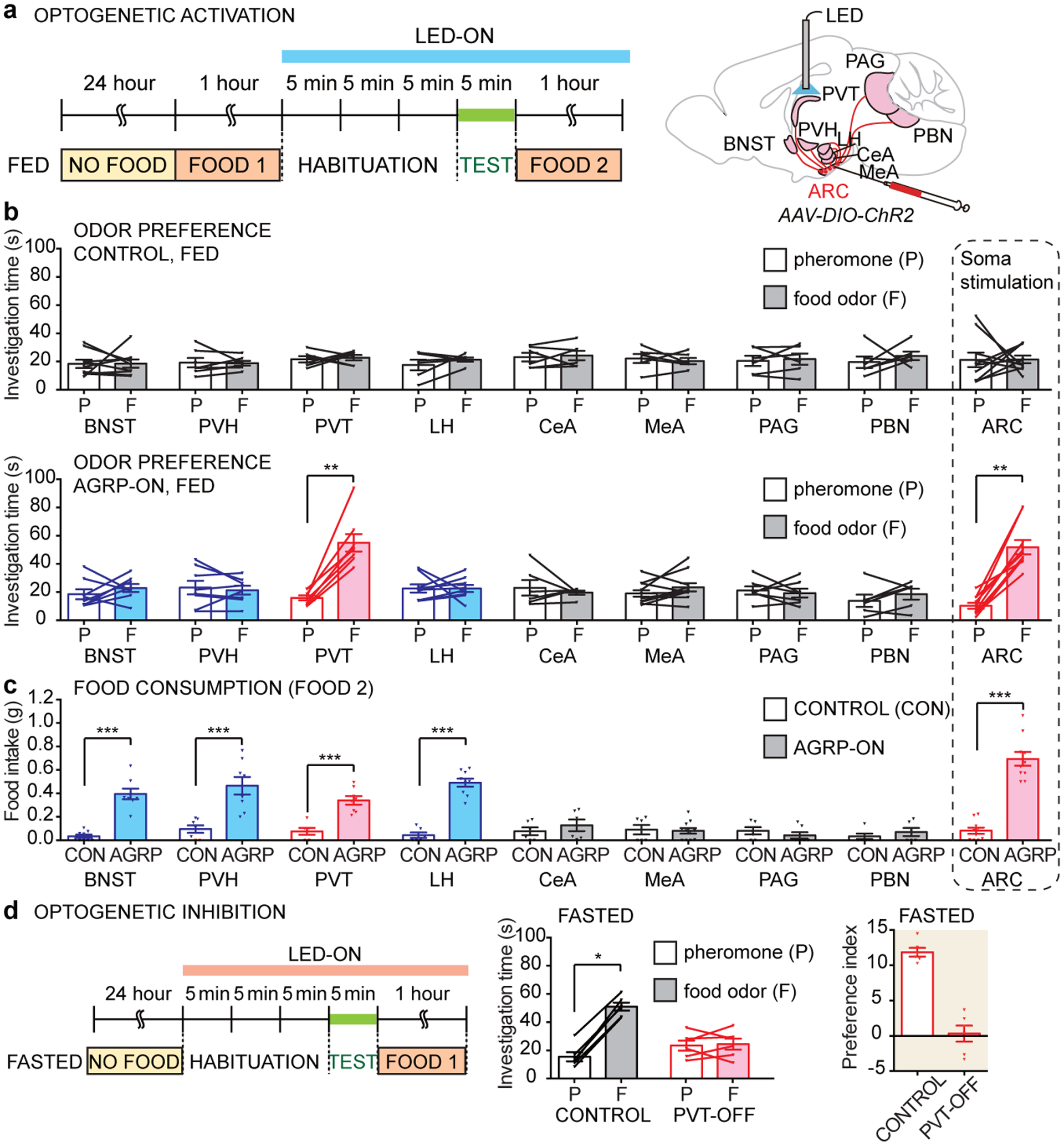
Internal state controls olfaction through poorly understood mechanisms. Odours that represent food, mates, competitors and predators activate parallel neural circuits that may be flexibly shaped by physiological need to alter behavioural outcome1. Here we identify a neuronal mechanism by which hunger selectively promotes attraction to food odours over other olfactory cues. Optogenetic activation of hypothalamic agouti-related peptide (AGRP) neurons enhances attraction to food odours but not to pheromones, and branch-specific activation and inhibition reveal a key role for projections to the paraventricular thalamus. Mice that lack neuropeptide Y (NPY) or NPY receptor type 5 (NPY5R) fail to prefer food odours over pheromones after fasting, and hunger-dependent food-odour attraction is restored by cell-specific NPY rescue in AGRP neurons. Furthermore, acute NPY injection immediately rescues food-odour preference without additional training, indicating that NPY is required for reading olfactory circuits during behavioural expression rather than writing olfactory circuits during odour learning. Together, these findings show that food-odour-responsive neurons comprise an olfactory subcircuit that listens to hunger state through thalamic NPY release, and more generally, provide mechanistic insights into how internal state regulates behaviour.
Conflict of interest statement
Competing interests declaration
The authors declare no competing financial interests.
Figures










References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
