

A novel p53 regulator, C16ORF72/TAPR1, buffers against telomerase inhibition
- PMID: 33660365
- PMCID: PMC8045932
- DOI: 10.1111/acel.13331
A novel p53 regulator, C16ORF72/TAPR1, buffers against telomerase inhibition
Abstract
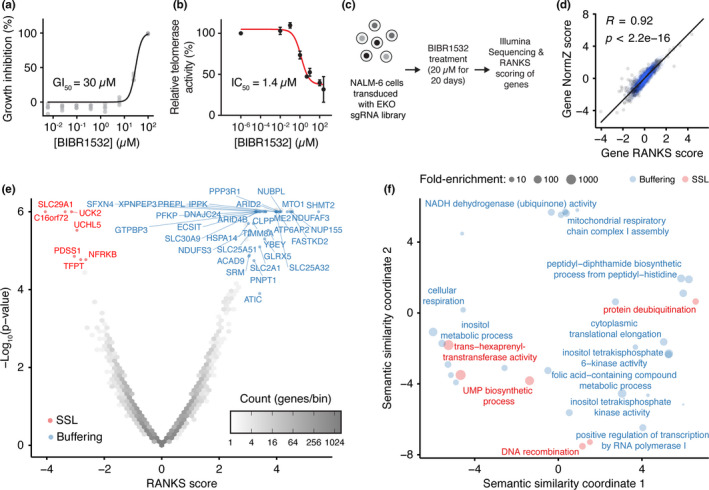
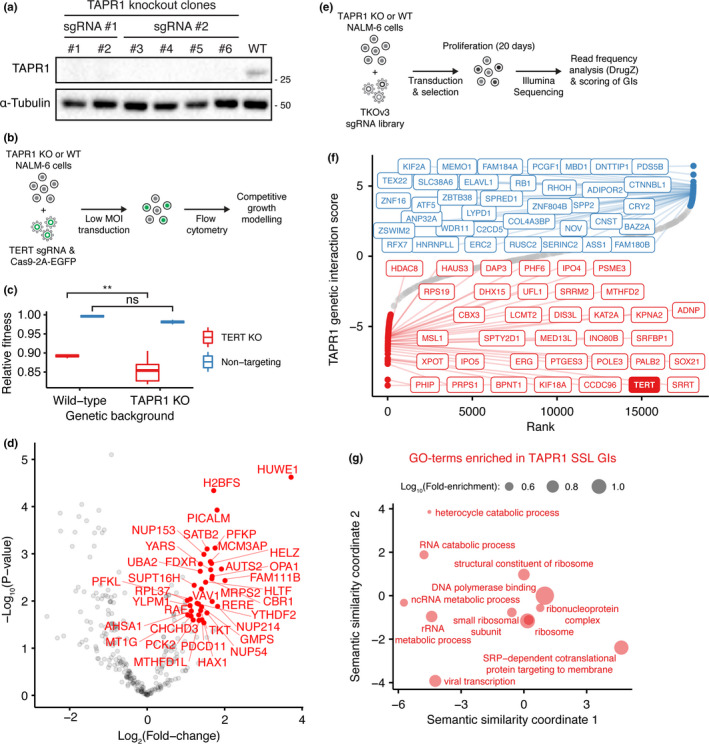
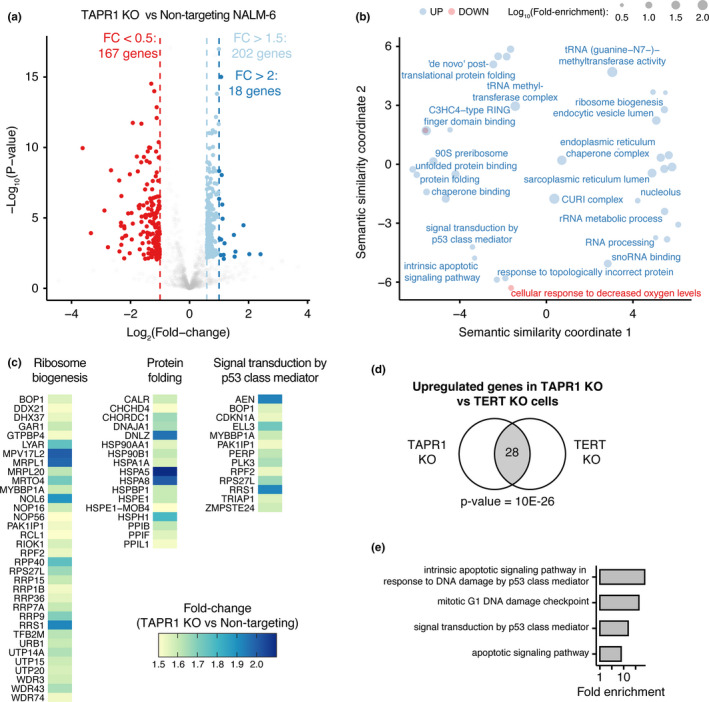
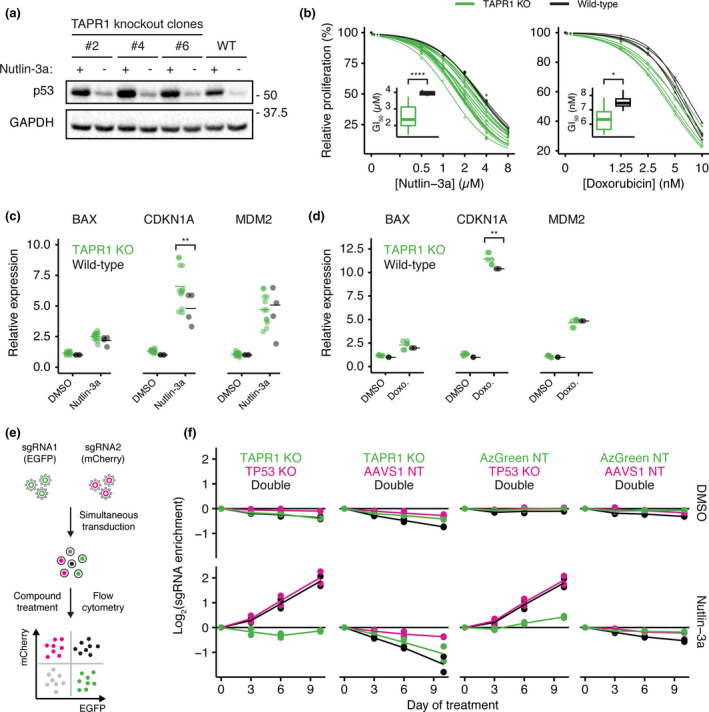
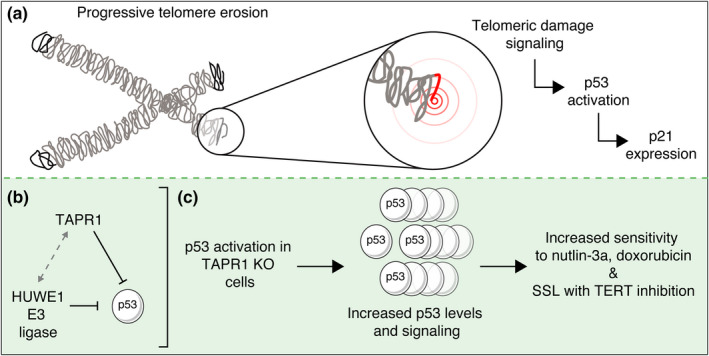
Telomere erosion in cells with insufficient levels of the telomerase reverse transcriptase (TERT), contributes to age-associated tissue dysfunction and senescence, and p53 plays a crucial role in this response. We undertook a genome-wide CRISPR screen to identify gene deletions that sensitized p53-positive human cells to telomerase inhibition. We uncovered a previously unannotated gene, C16ORF72, which we term Telomere Attrition and p53 Response 1 (TAPR1), that exhibited a synthetic-sick relationship with TERT loss. A subsequent genome-wide CRISPR screen in TAPR1-disrupted cells reciprocally identified TERT as a sensitizing gene deletion. Cells lacking TAPR1 or TERT possessed elevated p53 levels and transcriptional signatures consistent with p53 upregulation. The elevated p53 response in TERT- or TAPR1-deficient cells was exacerbated by treatment with the MDM2 inhibitor and p53 stabilizer nutlin-3a and coincided with a further reduction in cell fitness. Importantly, the sensitivity to treatment with nutlin-3a in TERT- or TAPR1-deficient cells was rescued by loss of p53. These data suggest that TAPR1 buffers against the deleterious consequences of telomere erosion or DNA damage by constraining p53. These findings identify C16ORF72/TAPR1 as new regulator at the nexus of telomere integrity and p53 regulation.
Keywords: C16ORF72; CRISPR-Cas9; Telomere Attrition and P53 Response 1; genome-wide screen; p53; synthetic-sick-lethal; telomerase inhibitor (BIBR1532); telomere erosion.
© 2021 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Benslimane, Y. , Bertomeu, T. , Coulombe‐Huntington, J. , McQuaid, M. , Sanchez‐Osuna, M. , Papadopoli, D. , Avizonis, D. , Russo, M. S. T. , Huard, C. , Topisirovic, I. , Wurtele, H. , Tyers, M. , & Harrington, L. (2020). Genome‐wide screens reveal that resveratrol induces replicative stress in human cells. Molecular Cell, 79(5), 846–856.e8. 10.1016/j.molcel.2020.07.010 - DOI - PubMed
-
- Bertomeu, T. , Coulombe‐Huntington, J. , Chatr‐Aryamontri, A. , Bourdages, K. G. , Coyaud, E. , Raught, B. , Xia, Y. , & Tyers, M. (2017). A high‐resolution genome‐wide CRISPR/Cas9 viability screen reveals structural features and contextual diversity of the human cell‐essential proteome. Molecular and Cellular Biology, 38(1), e00302‐17. 10.1128/MCB.00302-17 - DOI - PMC - PubMed
-
- Brennan, C. M. , Vaites, L. P. , Wells, J. N. , Santaguida, S. , Paulo, J. A. , Storchova, Z. , Harper, J. W. , Marsh, J. A. , & Amon, A. (2019). Protein aggregation mediates stoichiometry of protein complexes in aneuploid cells. Genes & Development, 33(15–16), 1031–1047. 10.1101/gad.327494.119 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
