

Is γ-secretase a beneficial inactivating enzyme of the toxic APP C-terminal fragment C99?
- PMID: 33662398
- PMCID: PMC8027268
- DOI: 10.1016/j.jbc.2021.100489
Is γ-secretase a beneficial inactivating enzyme of the toxic APP C-terminal fragment C99?
Abstract
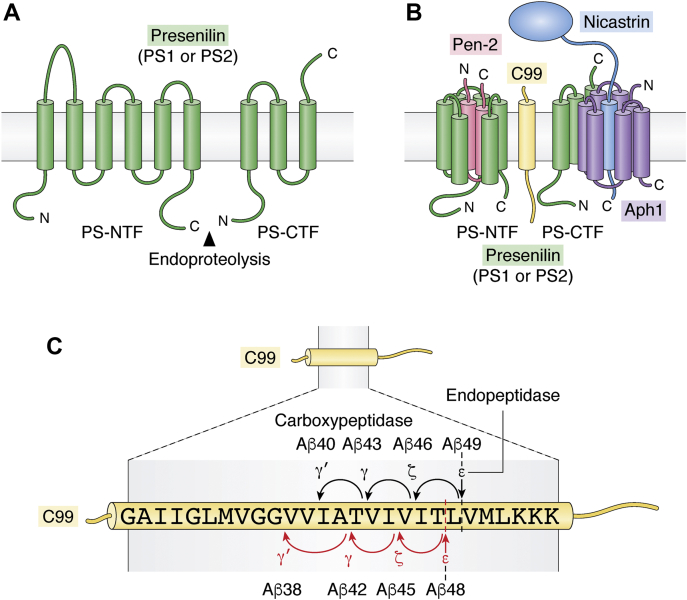
Genetic, biochemical, and anatomical grounds led to the proposal of the amyloid cascade hypothesis centered on the accumulation of amyloid beta peptides (Aβ) to explain Alzheimer's disease (AD) etiology. In this context, a bulk of efforts have aimed at developing therapeutic strategies seeking to reduce Aβ levels, either by blocking its production (γ- and β-secretase inhibitors) or by neutralizing it once formed (Aβ-directed immunotherapies). However, so far the vast majority of, if not all, clinical trials based on these strategies have failed, since they have not been able to restore cognitive function in AD patients, and even in many cases, they have worsened the clinical picture. We here propose that AD could be more complex than a simple Aβ-linked pathology and discuss the possibility that a way to reconcile undoubted genetic evidences linking processing of APP to AD and a consistent failure of Aβ-based clinical trials could be to envision the pathological contribution of the direct precursor of Aβ, the β-secretase-derived C-terminal fragment of APP, βCTF, also referred to as C99. In this review, we summarize scientific evidences pointing to C99 as an early contributor to AD and postulate that γ-secretase should be considered as not only an Aβ-generating protease, but also a beneficial C99-inactivating enzyme. In that sense, we discuss the limitations of molecules targeting γ-secretase and propose alternative strategies seeking to reduce C99 levels by other means and notably by enhancing its lysosomal degradation.
Keywords: APP C-terminal fragments; C99; clinical trials; toxicity; β-secretase; γ-secretase.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



References
-
- Calderon-Garciduenas A.L., Duyckaerts C. Alzheimer disease. Handbook Clin. Neurol. 2017;145:325–337. - PubMed
-
- Sengoku R. Aging and Alzheimer's disease pathology. Neuropathology. 2020;40:22–29. - PubMed
-
- Glenner G.G., Wong C.W. Alzheimer's disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. BBRC. 1984;120:885–890. - PubMed
-
- Brion J.P., Flament-Durand J., Dustin P. Alzheimer's disease and tau proteins. Lancet. 1986;2:1098. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
