

Heterozygous APC germline mutations impart predisposition to colorectal cancer
- PMID: 33664379
- PMCID: PMC7933349
- DOI: 10.1038/s41598-021-84564-4
Heterozygous APC germline mutations impart predisposition to colorectal cancer
Abstract
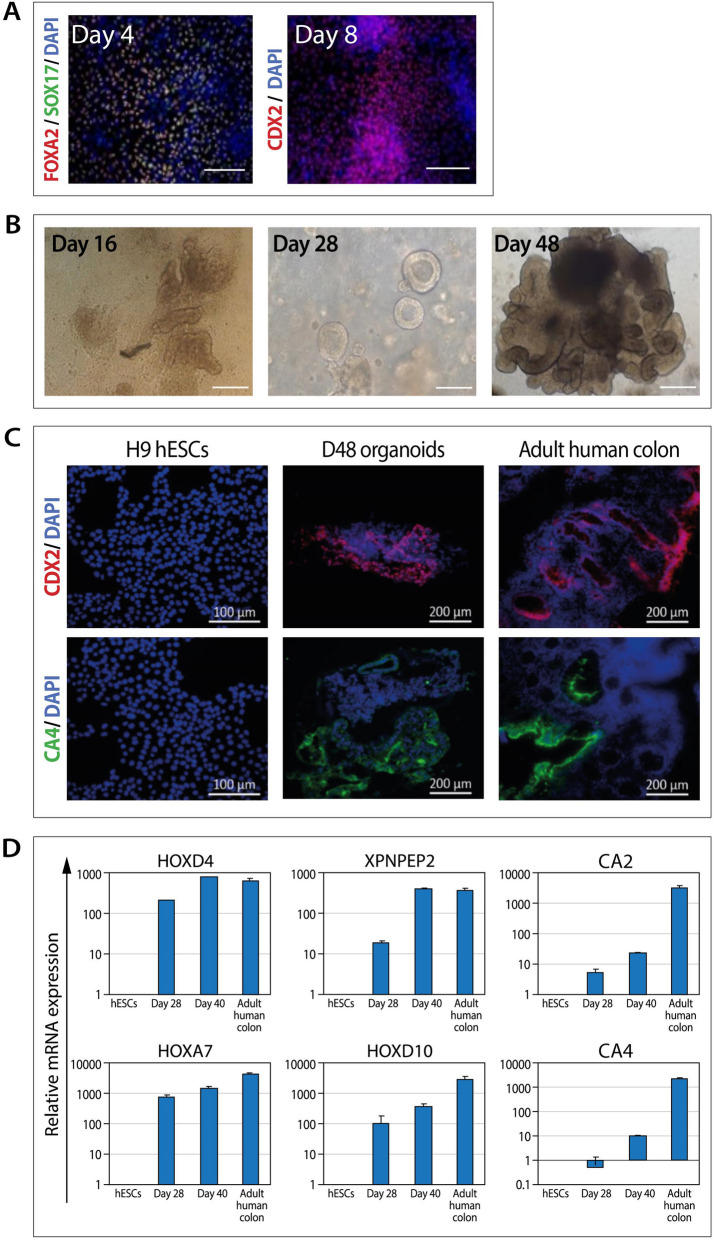
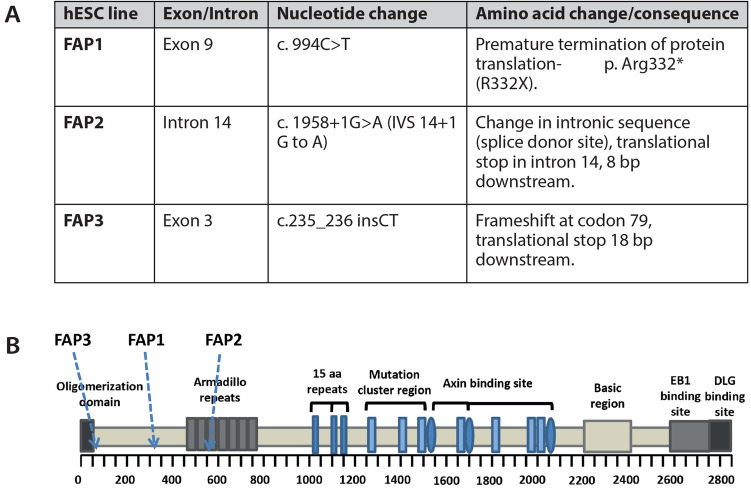
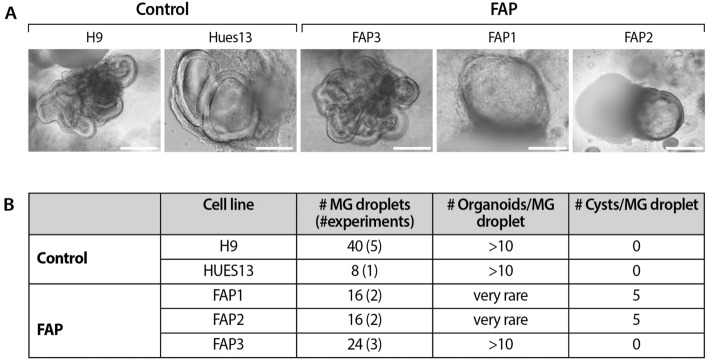
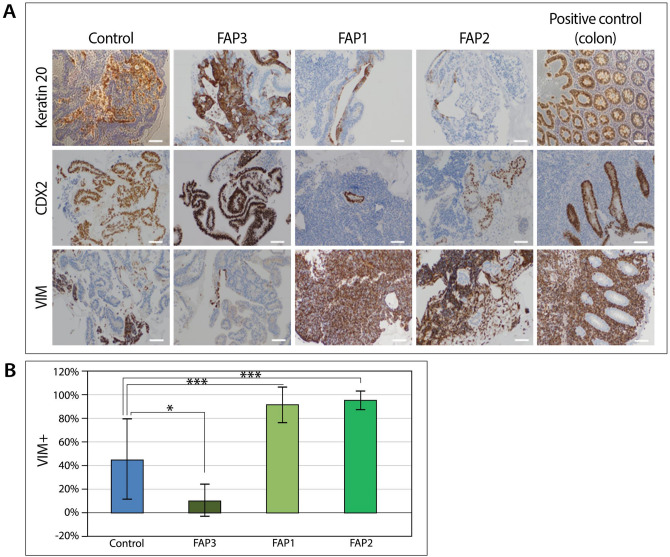
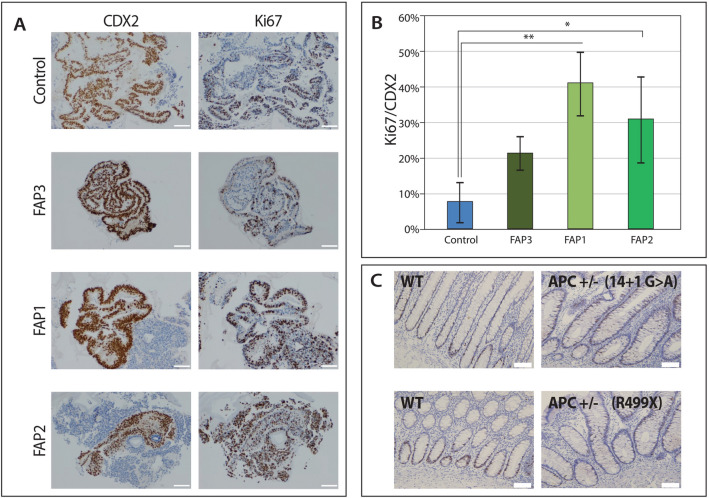
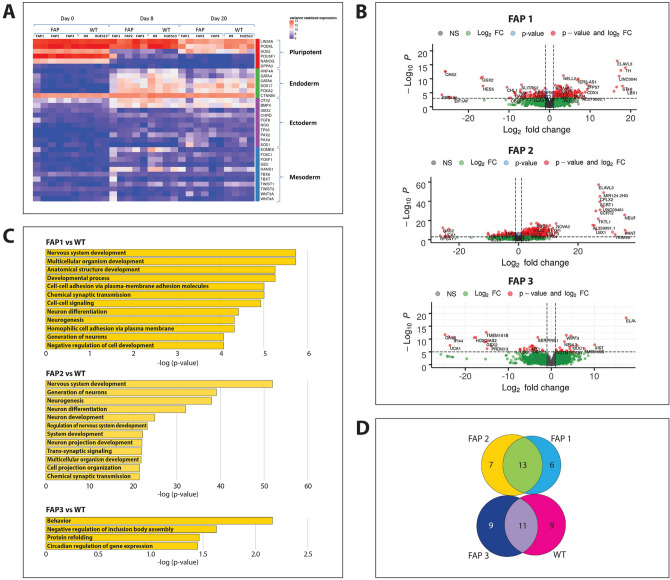
Familial adenomatous polyposis (FAP) is an inherited syndrome caused by a heterozygous adenomatous polyposis coli (APC) germline mutation, associated with a profound lifetime risk for colorectal cancer. While it is well accepted that tumorigenic transformation is initiated following acquisition of a second mutation and loss of function of the APC gene, the role of heterozygous APC mutation in this process is yet to be discovered. This work aimed to explore whether a heterozygous APC mutation induces molecular defects underlying tumorigenic transformation and how different APC germline mutations predict disease severity. Three FAP-human embryonic stem cell lines (FAP1/2/3-hESC lines) carrying germline mutations at different locations of the APC gene, and two control hESC lines free of the APC mutation, were differentiated into colon organoids and analyzed by immunohistochemistry and RNA sequencing. In addition, data regarding the genotype and clinical phenotype of the embryo donor parents were collected from medical records. FAP-hESCs carrying a complete loss-of-function of a single APC allele (FAP3) generated complex and molecularly mature colon organoids, which were similar to controls. In contrast, FAP-hESCs carrying APC truncation mutations (FAP1 and FAP2) generated only few cyst-like structures and cell aggregates of various shape, occasionally with luminal parts, which aligned with their failure to upregulate critical differentiation genes early in the process, as shown by RNA sequencing. Abnormal disease phenotype was shown also in non-pathological colon of FAP patients by the randomly distribution of proliferating cells throughout the crypts, compared to their focused localization in the lower part of the crypt in healthy/non-FAP patients. Genotype/phenotype analysis revealed correlations between the colon organoid maturation potential and FAP severity in the carrier parents. In conclusion, this study suggest that a single truncated APC allele is sufficient to initiate early molecular tumorigenic activity. In addition, the results hint that patient-specific hESC-derived colon organoids can probably predict disease severity among FAP patients.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The effect of a germline mutation in the APC gene on β-catenin in human embryonic stem cells.BMC Cancer. 2016 Dec 23;16(1):952. doi: 10.1186/s12885-016-2809-9. BMC Cancer. 2016. PMID: 28010732 Free PMC article.
-
A neoepitope derived from a novel human germline APC gene mutation in familial adenomatous polyposis shows selective immunogenicity.PLoS One. 2018 Sep 26;13(9):e0203845. doi: 10.1371/journal.pone.0203845. eCollection 2018. PLoS One. 2018. PMID: 30256815 Free PMC article.
-
Predicting colorectal cancer risk in FAP patients using patient-specific organoids.Cancer Gene Ther. 2025 Sep;32(9):997-1007. doi: 10.1038/s41417-025-00923-7. Epub 2025 Jul 22. Cancer Gene Ther. 2025. PMID: 40695959 Free PMC article.
-
Correlations between mutation site in APC and phenotype of familial adenomatous polyposis (FAP): a review of the literature.Crit Rev Oncol Hematol. 2007 Feb;61(2):153-61. doi: 10.1016/j.critrevonc.2006.07.004. Epub 2006 Oct 24. Crit Rev Oncol Hematol. 2007. PMID: 17064931 Review.
-
The genetics of familial adenomatous polyposis (FAP) and MutYH-associated polyposis (MAP).Acta Gastroenterol Belg. 2011 Sep;74(3):421-6. Acta Gastroenterol Belg. 2011. PMID: 22103048 Review.
Cited by
-
The Physiological Function and Potential Role of the Ubiquitous Na+/H+ Exchanger Isoform 8 (NHE8): An Overview Data.Int J Mol Sci. 2022 Sep 17;23(18):10857. doi: 10.3390/ijms231810857. Int J Mol Sci. 2022. PMID: 36142772 Free PMC article. Review.
-
Colorectal Cancer, Liver Metastases and Biotherapies.Biomedicines. 2021 Jul 26;9(8):894. doi: 10.3390/biomedicines9080894. Biomedicines. 2021. PMID: 34440099 Free PMC article. Review.
-
Altered lipid metabolism in APC-driven colorectal cancer: the potential for therapeutic intervention.Front Oncol. 2024 Mar 25;14:1343061. doi: 10.3389/fonc.2024.1343061. eCollection 2024. Front Oncol. 2024. PMID: 38590663 Free PMC article. Review.
-
Rapamycin rescues APC-mutated colon organoid differentiation.Cancer Gene Ther. 2025 Sep;32(9):1008-1017. doi: 10.1038/s41417-025-00935-3. Epub 2025 Jul 23. Cancer Gene Ther. 2025. PMID: 40702333 Free PMC article.
-
APC-driven actin nucleation powers collective cell dynamics in colorectal cancer cells.iScience. 2023 Apr 6;26(5):106583. doi: 10.1016/j.isci.2023.106583. eCollection 2023 May 19. iScience. 2023. PMID: 37128612 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
