

Granzyme A inhibition reduces inflammation and increases survival during abdominal sepsis
- PMID: 33664861
- PMCID: PMC7914344
- DOI: 10.7150/thno.49288
Granzyme A inhibition reduces inflammation and increases survival during abdominal sepsis
Abstract
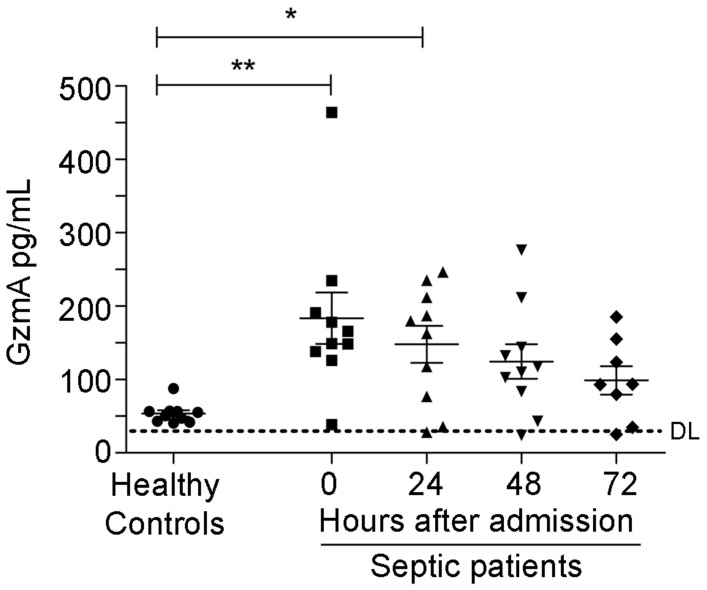
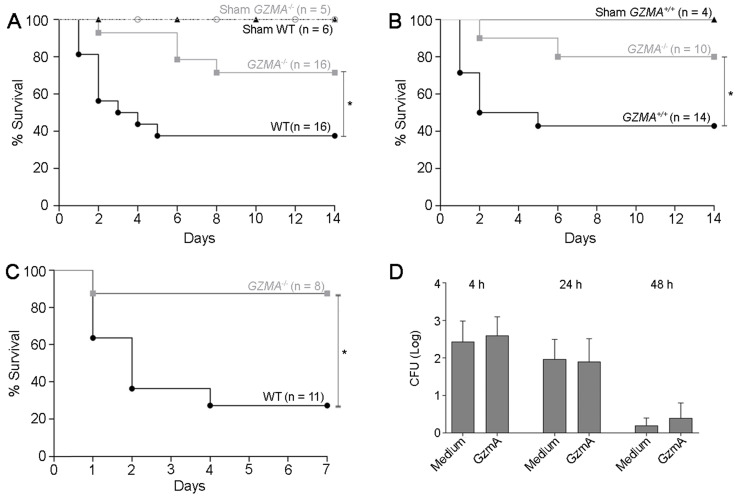
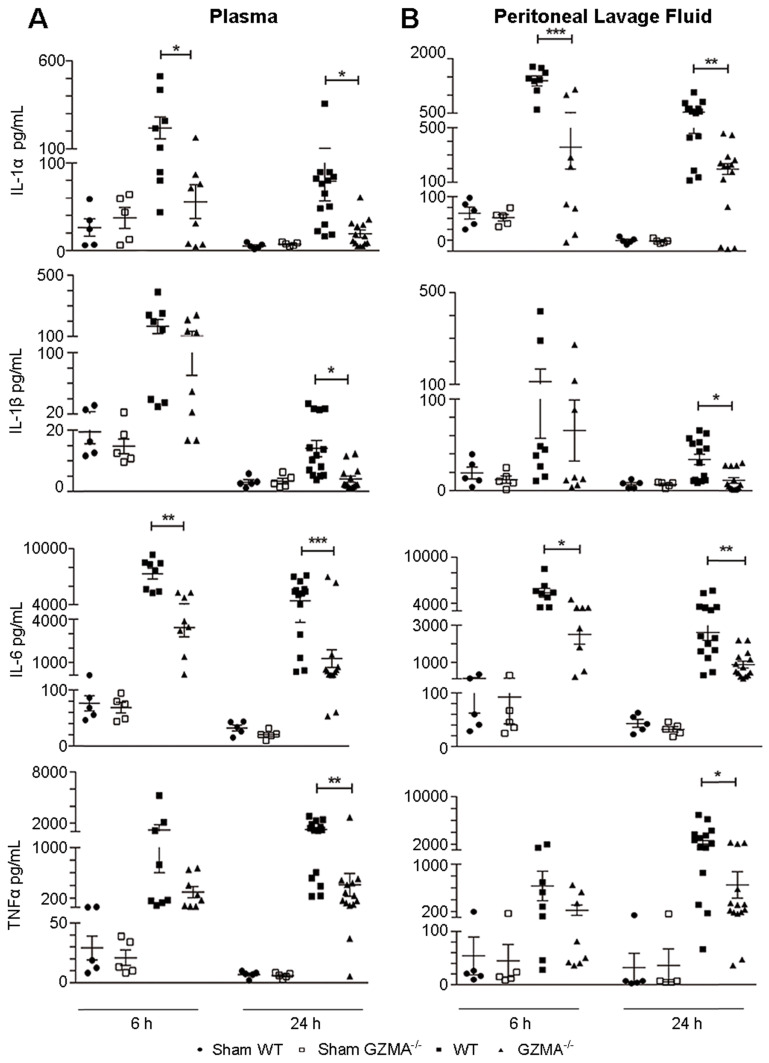
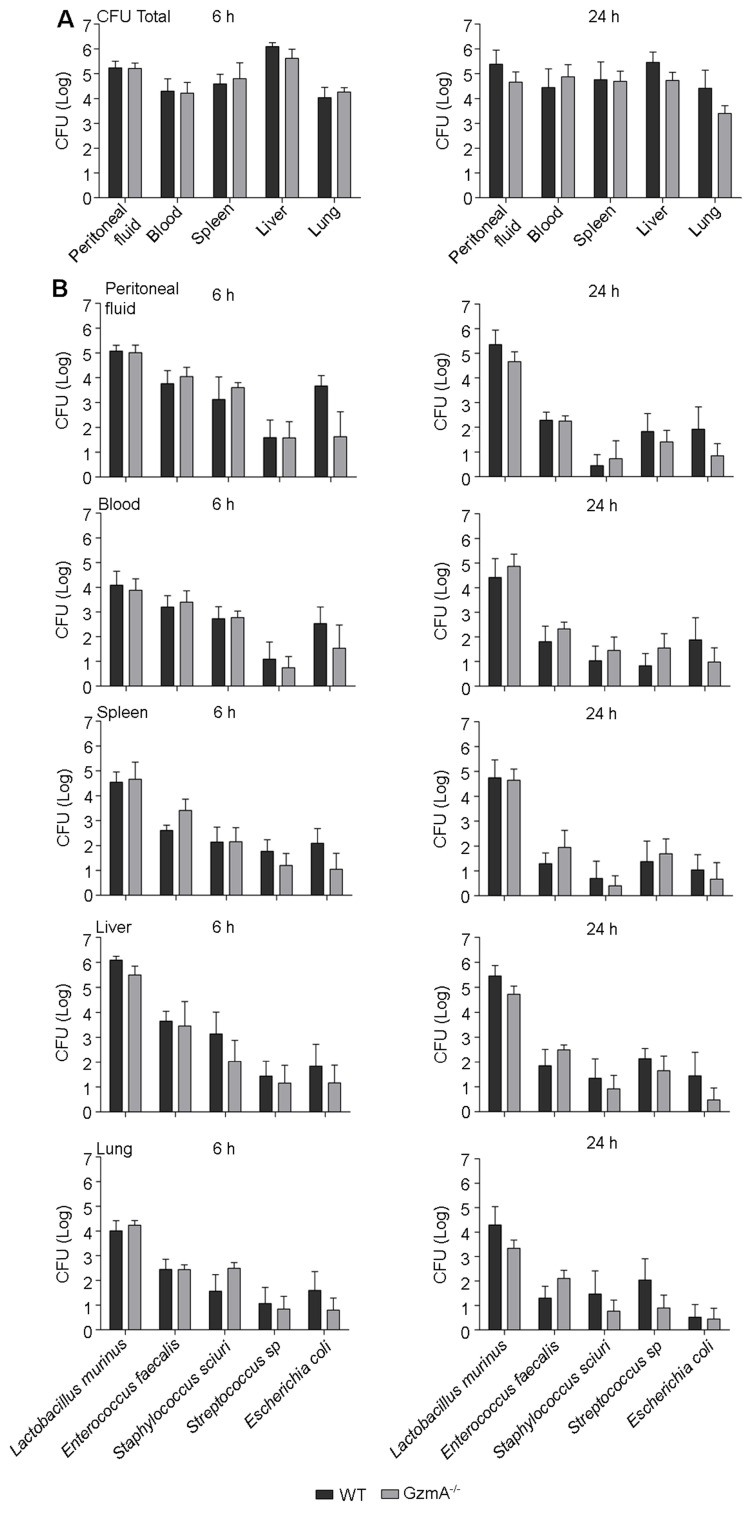
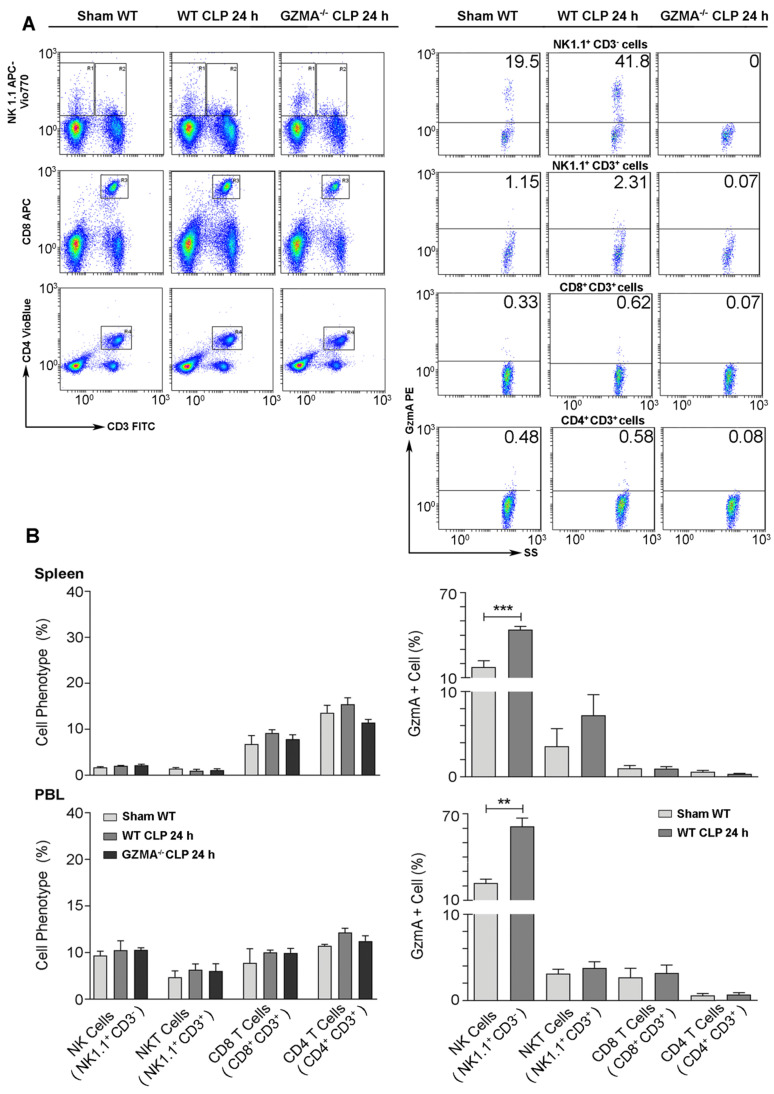
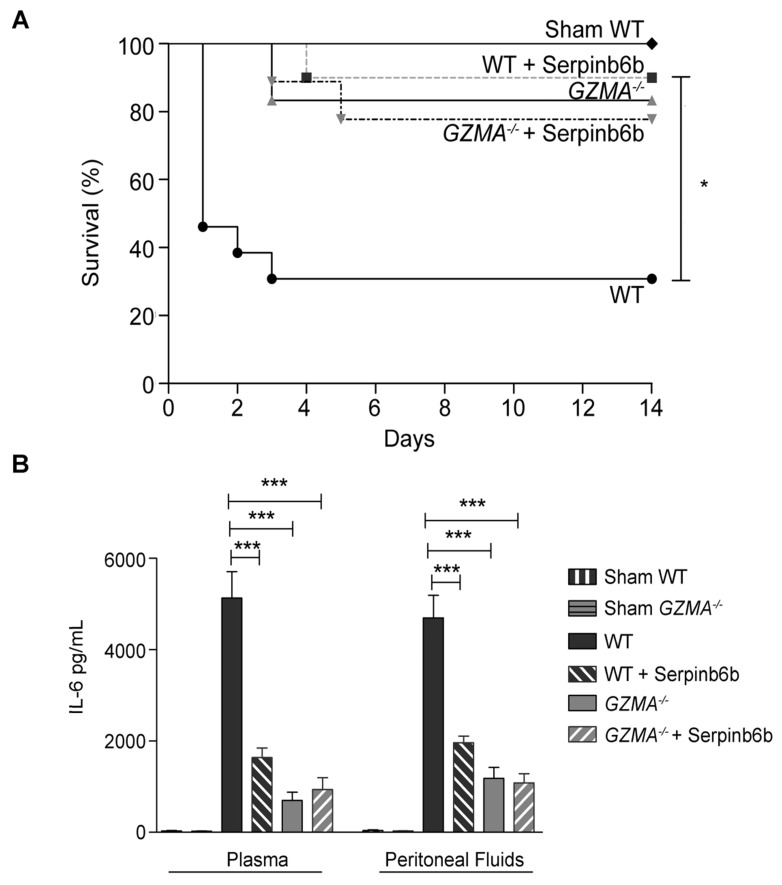
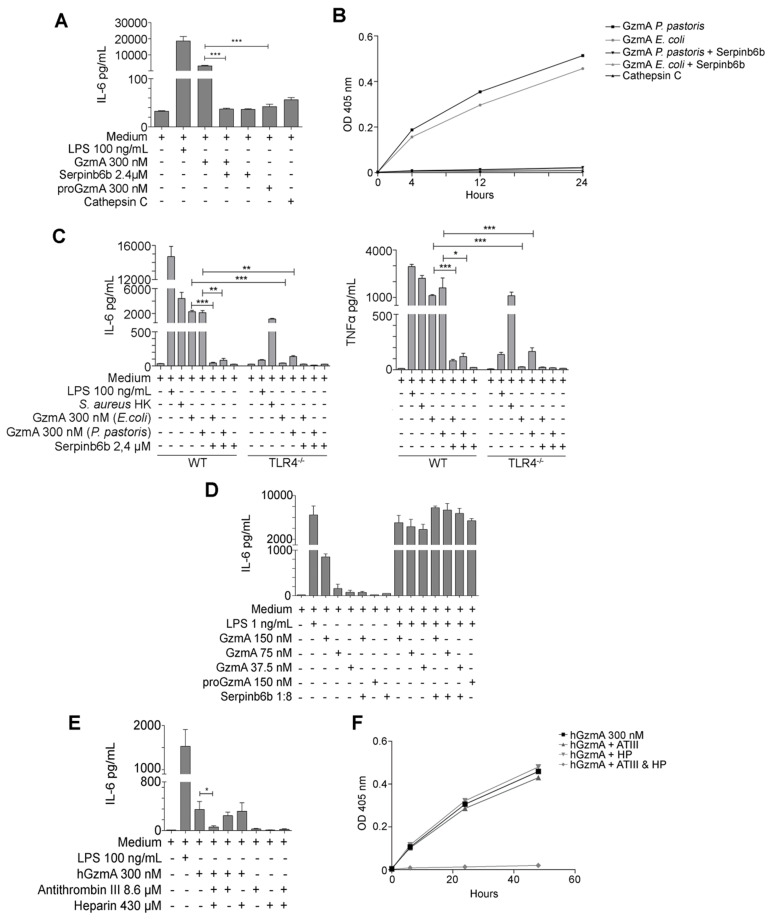
Aims: Peritonitis is one of the most common causes of sepsis, a serious syndrome characterized by a dysregulated systemic inflammatory response. Recent evidence suggests that Granzyme A (GzmA), a serine protease mainly expressed by NK and T cells, could act as a proinflammatory mediator and could play an important role in the pathogenesis of sepsis. This work aims to analyze the role and the therapeutic potential of GzmA in the pathogenesis of peritoneal sepsis. Methods: The level of extracellular GzmA as well as GzmA activity were analyzed in serum from healthy volunteers and patients with confirmed peritonitis and were correlated with the Sequential Organ Failure Assessment (SOFA) score. Peritonitis was induced in C57Bl/6 (WT) and GzmA-/- mice by cecal ligation and puncture (CLP). Mice were treated intraperitoneally with antibiotics alone or in combination serpinb6b, a specific GzmA inhibitor, for 5 days. Mouse survival was monitored during 14 days, levels of some proinflammatory cytokines were measured in serum and bacterial load and diversity was analyzed in blood and spleen at different times. Results: Clinically, elevated GzmA was observed in serum from patients with abdominal sepsis suggesting that GzmA plays an important role in this pathology. In the CLP model GzmA deficient mice, or WT mice treated with an extracellular GzmA inhibitor, showed increased survival, which correlated with a reduction in proinflammatory markers in both serum and peritoneal lavage fluid. GzmA deficiency did not influence bacterial load in blood and spleen and GzmA did not affect bacterial replication in macrophages in vitro, indicating that GzmA has no role in bacterial control. Analysis of GzmA in lymphoid cells following CLP showed that it was mainly expressed by NK cells. Mechanistically, we found that extracellular active GzmA acts as a proinflammatory mediator in macrophages by inducing the TLR4-dependent expression of IL-6 and TNFα. Conclusions: Our findings implicate GzmA as a key regulator of the inflammatory response during abdominal sepsis and provide solid evidences about its therapeutic potential for the treatment of this severe pathology.
Keywords: Granzyme A; cecal ligation and puncture.; inflammation; peritonitis; sepsis.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures
References
-
- Brown D, Caballero Alvarado JA. Sepsis (Septic), Peritonitis. StatPearls. Treasure Island (FL) 2019. - PubMed
-
- Gotts JE, Matthay MA. Sepsis: pathophysiology and clinical management. Bmj. 2016;353:i1585. - PubMed
-
- Arias M, Martinez-Lostao L, Santiago L, Ferrandez A, Granville DJ, Pardo J. The Untold Story of Granzymes in Oncoimmunology: Novel Opportunities with Old Acquaintances. Trends in cancer. 2017;3:407–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
