

Role of Constitutive STAR in Leydig Cells
- PMID: 33670702
- PMCID: PMC7922663
- DOI: 10.3390/ijms22042021
Role of Constitutive STAR in Leydig Cells
Abstract
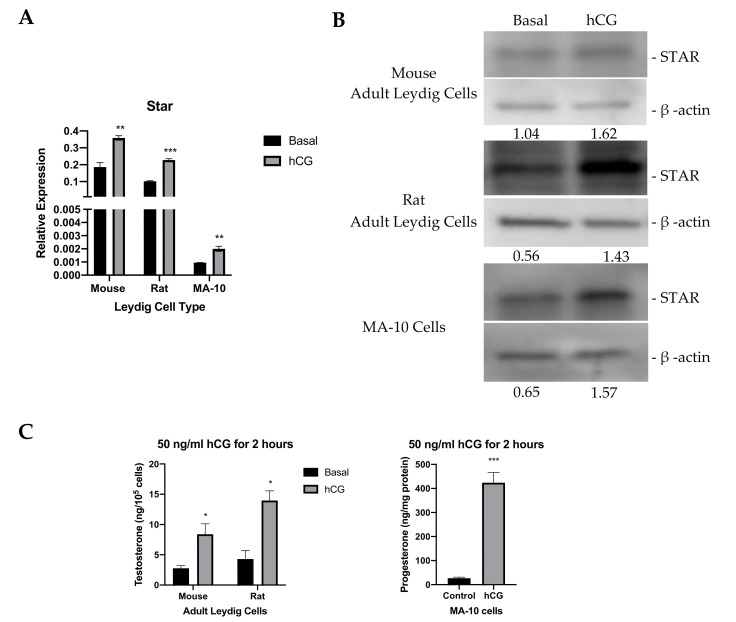
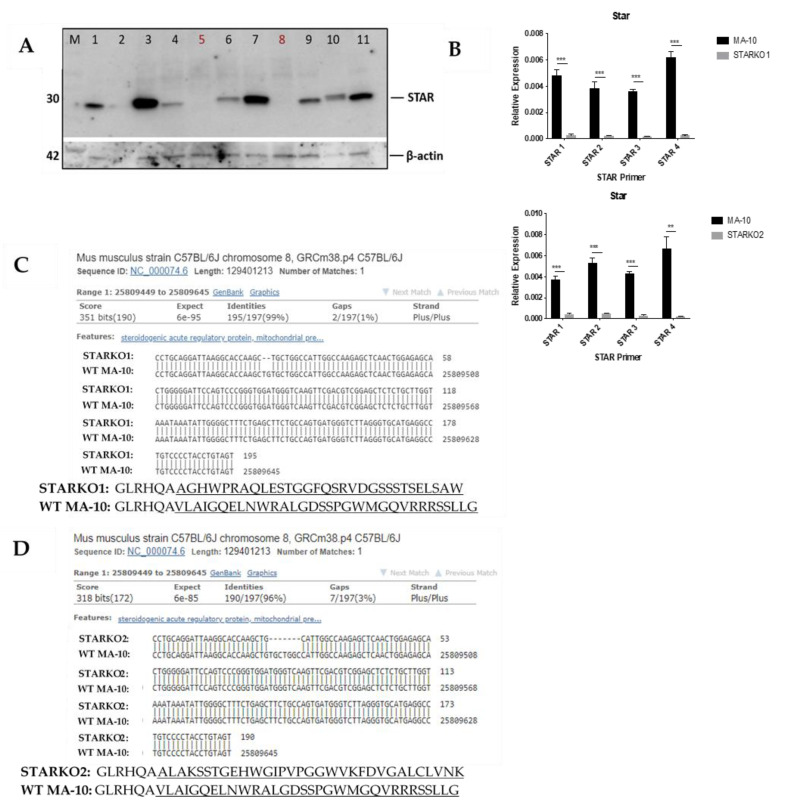
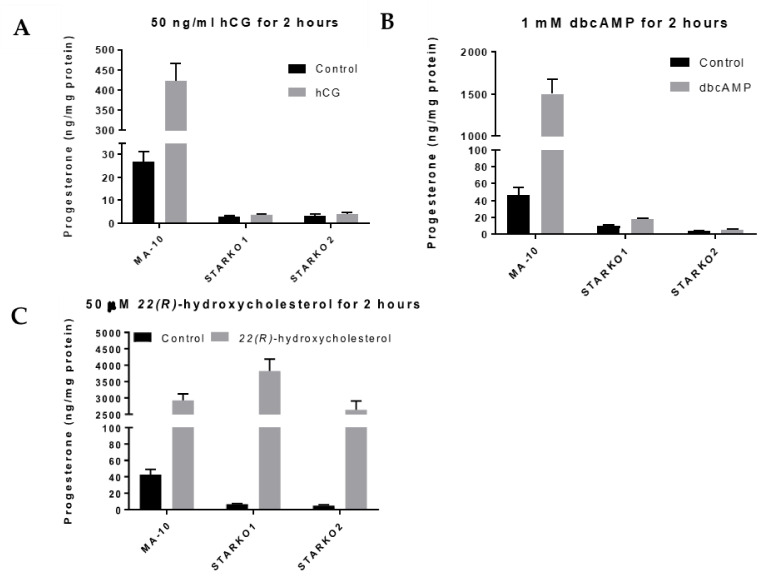
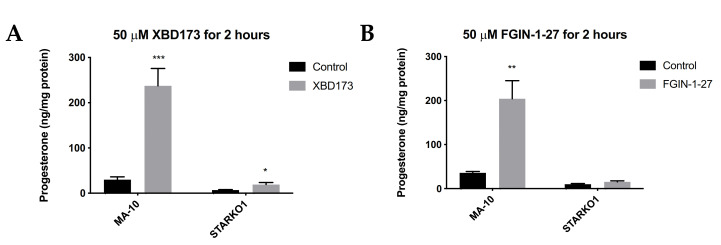
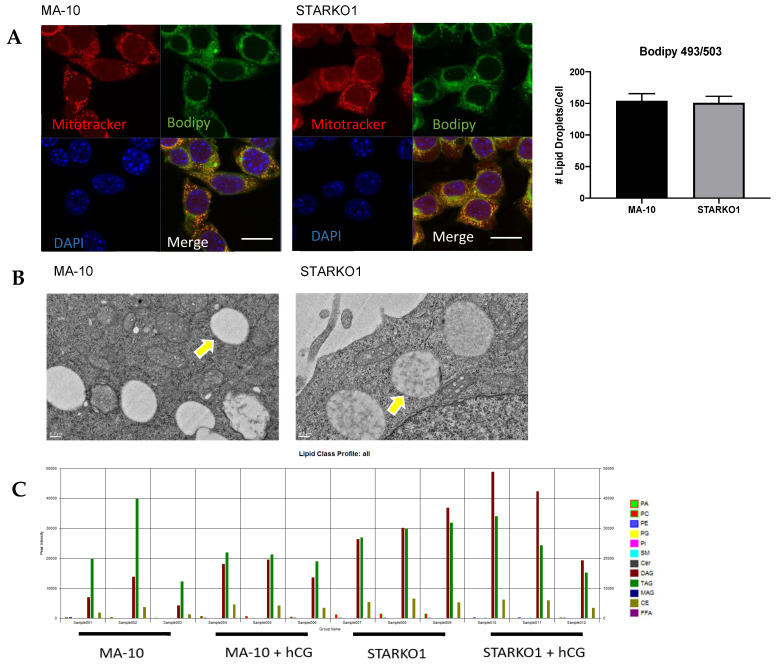
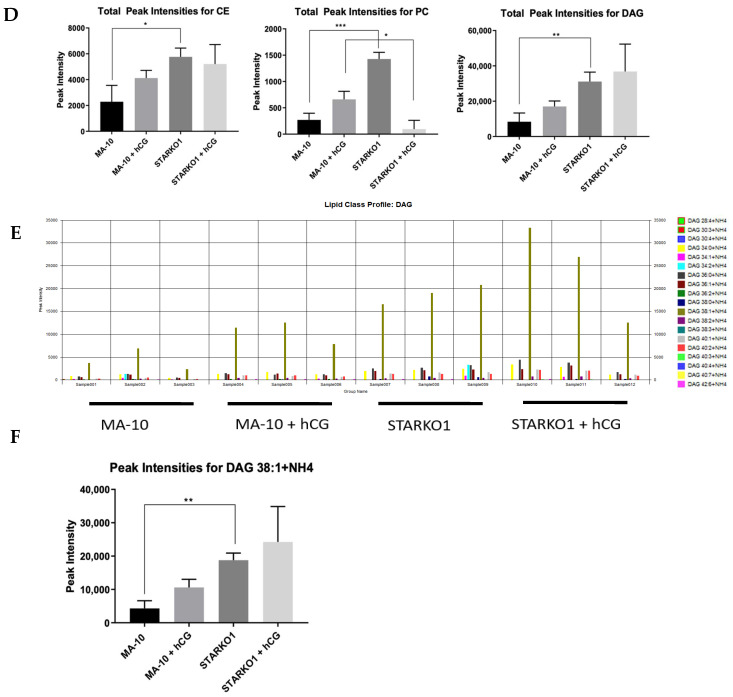
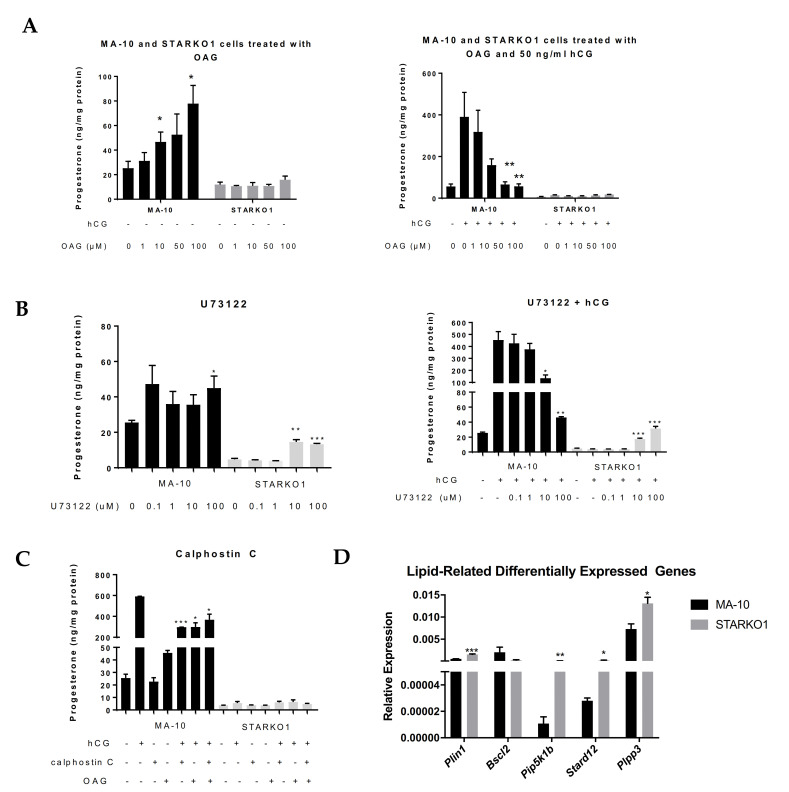
Leydig cells contain significant amounts of constitutively produced steroidogenic acute regulatory protein (STAR; STARD1). Hormone-induced STAR plays an essential role in inducing the transfer of cholesterol into the mitochondria for hormone-dependent steroidogenesis. STAR acts at the outer mitochondrial membrane, where it interacts with a protein complex, which includes the translocator protein (TSPO). Mutations in STAR cause lipoid congenital adrenal hyperplasia (lipoid CAH), a disorder characterized by severe defects in adrenal and gonadal steroid production; in Leydig cells, the defects are seen mainly after the onset of hormone-dependent androgen formation. The function of constitutive STAR in Leydig cells is unknown. We generated STAR knockout (KO) MA-10 mouse tumor Leydig cells and showed that STAR KO cells failed to form progesterone in response to dibutyryl-cAMP and to TSPO drug ligands, but not to 22(R)-hydroxycholesterol, which is a membrane-permeable intermediate of the CYP11A1 reaction. Electron microscopy of STAR KO cells revealed that the number and size of lipid droplets were similar to those in wild-type (WT) MA-10 cells. However, the density of lipid droplets in STAR KO cells was drastically different than that seen in WT cells. We isolated the lipid droplets and analyzed their content by liquid chromatography-mass spectrometry. There was a significant increase in cholesteryl ester and phosphatidylcholine content in STAR KO cell lipid droplets, but the most abundant increase was in the amount of diacylglycerol (DAG); DAG 38:1 was the predominantly affected species. Lastly, we identified genes involved in DAG signaling and lipid metabolism which were differentially expressed between WT MA-10 and STAR KO cells. These results suggest that constitutive STAR in Leydig cells is involved in DAG accumulation in lipid droplets, in addition to cholesterol transport. The former event may affect cell functions mediated by DAG signaling.
Keywords: STAR; cholesterol; diacylglycerol; lipid droplets; steroidogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Rone M.B., Midzak A.S., Issop L., Rammouz G., Jagannathan S., Fan J., Papadopoulos V. Identification of a dynamic mitochondrial protein complex driving cholesterol import, trafficking, and metabolism to steroid hormones. Mol. Endocrinol. 2012;26:1868–1882. doi: 10.1210/me.2012-1159. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
