

Oligomeric Aβ1-42 Induces an AMD-Like Phenotype and Accumulates in Lysosomes to Impair RPE Function
- PMID: 33671133
- PMCID: PMC7922851
- DOI: 10.3390/cells10020413
Oligomeric Aβ1-42 Induces an AMD-Like Phenotype and Accumulates in Lysosomes to Impair RPE Function
Abstract
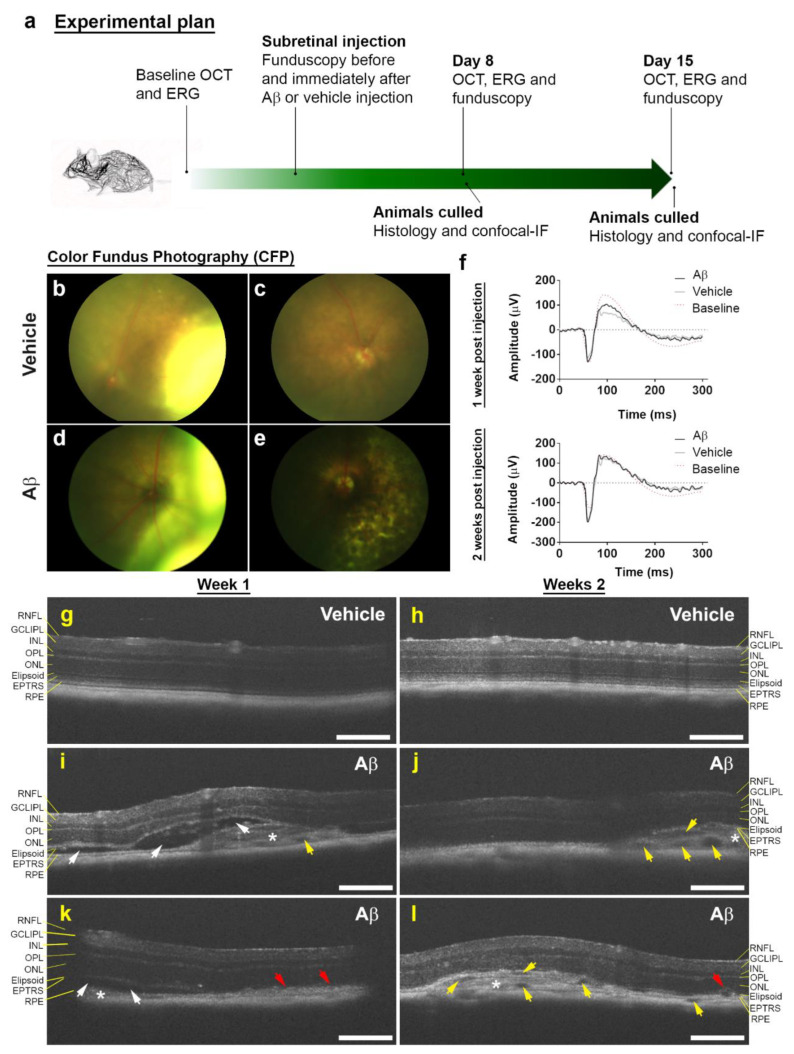
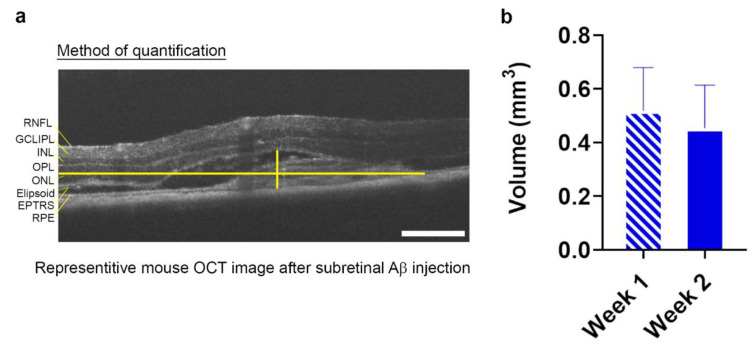
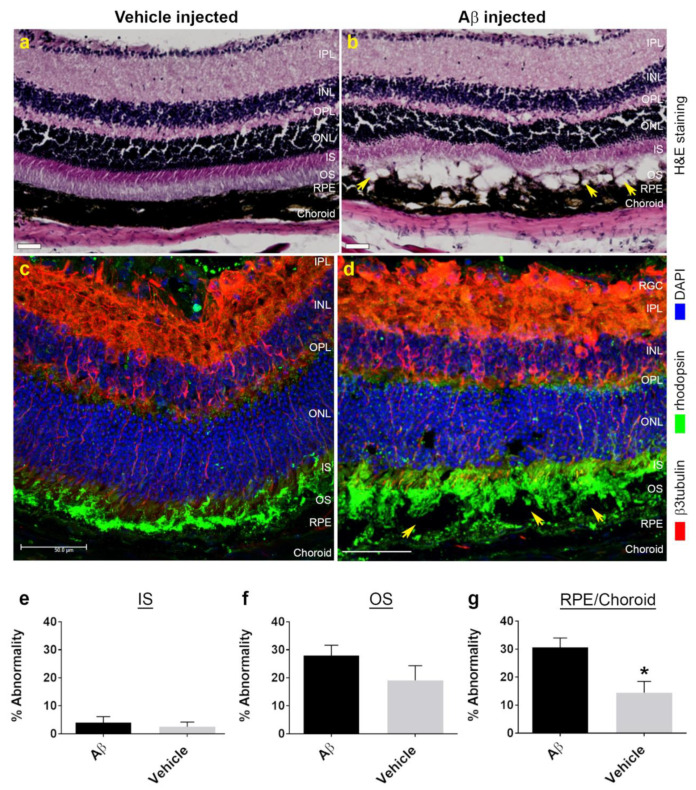
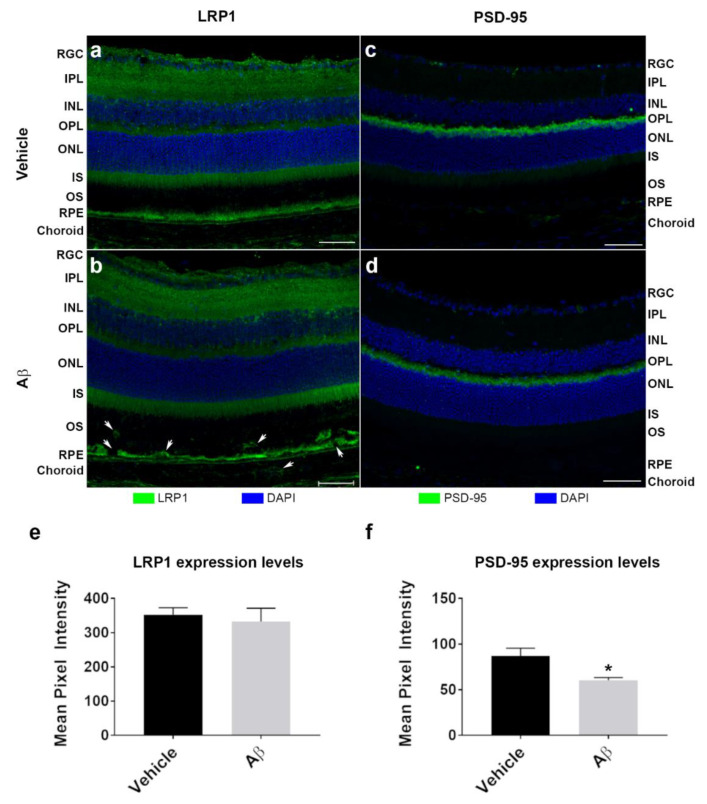
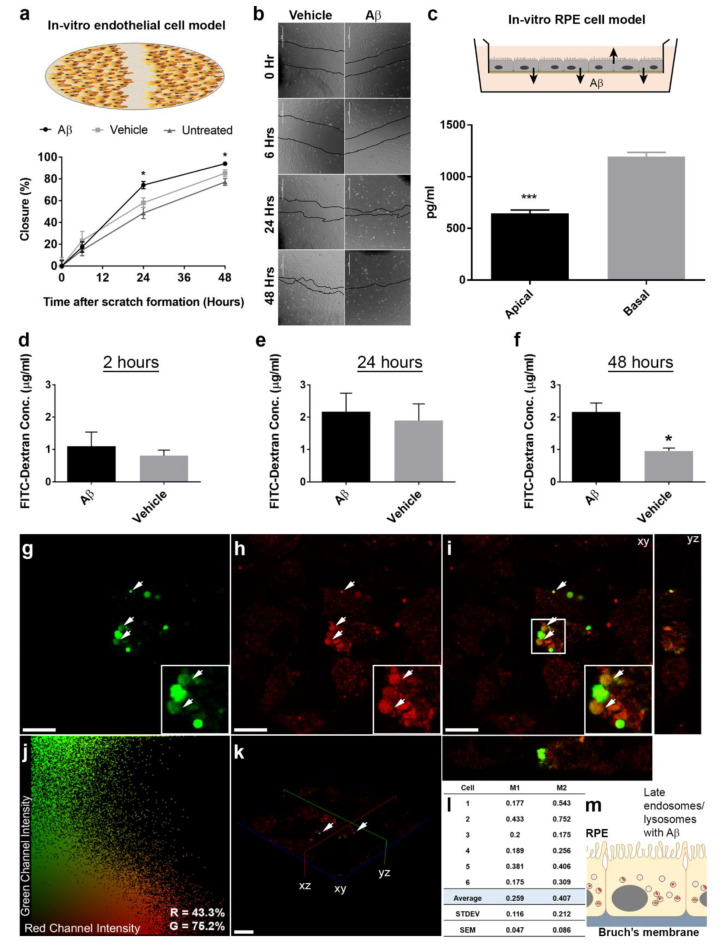
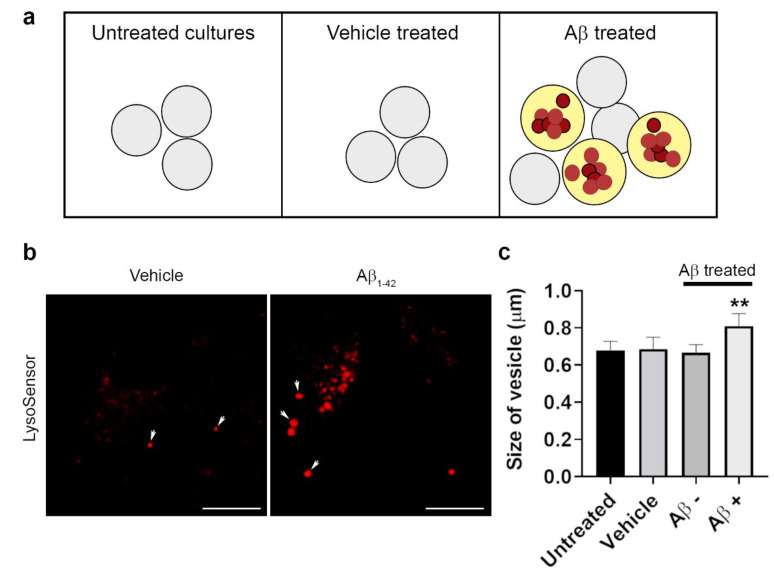
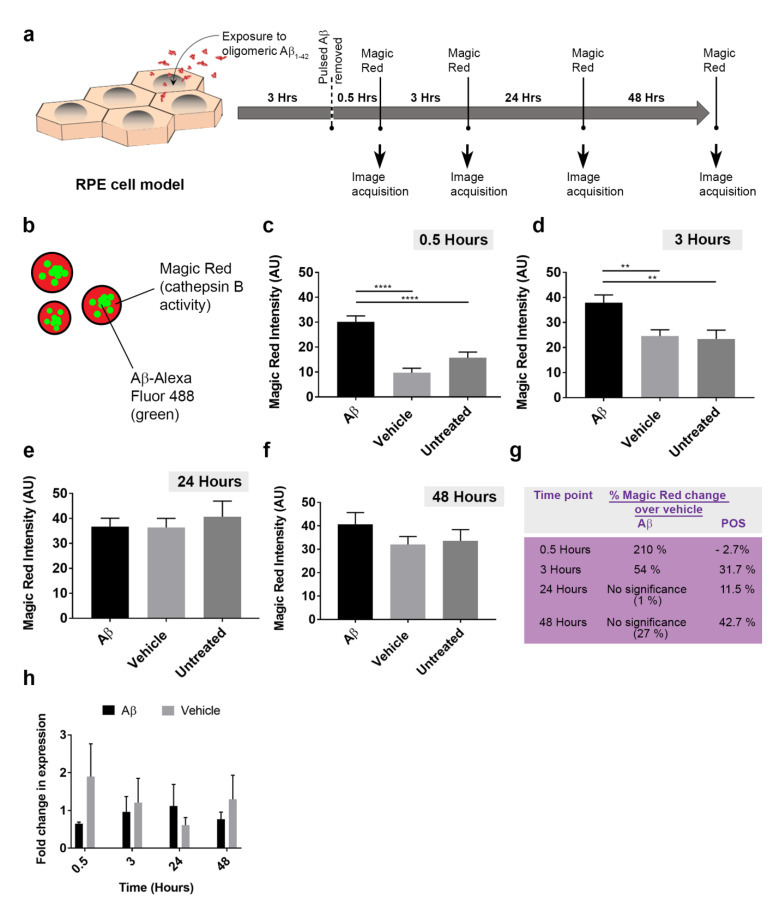
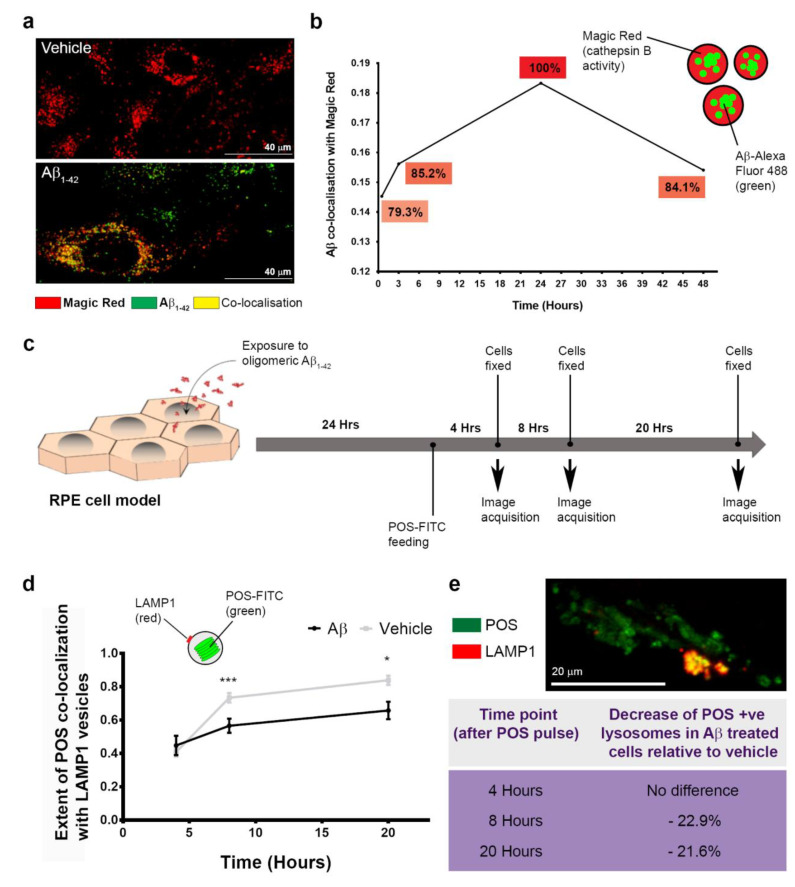
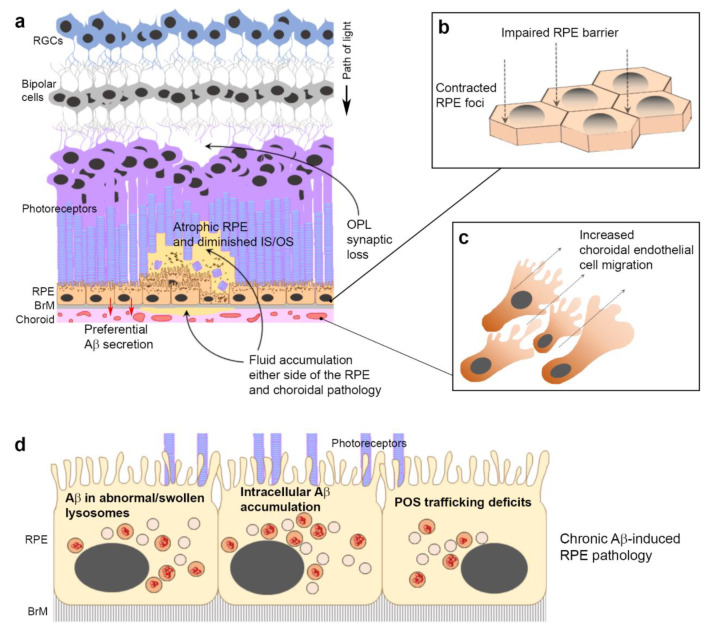
Alzheimer's disease-associated amyloid beta (Aβ) proteins accumulate in the outer retina with increasing age and in eyes of age-related macular degeneration (AMD) patients. To study Aβ-induced retinopathy, wild-type mice were injected with nanomolar human oligomeric Aβ1-42, which recapitulate the Aβ burden reported in human donor eyes. In vitro studies investigated the cellular effects of Aβ in endothelial and retinal pigment epithelial (RPE) cells. Results show subretinal Aβ-induced focal AMD-like pathology within 2 weeks. Aβ exposure caused endothelial cell migration, and morphological and barrier alterations to the RPE. Aβ co-localized to late-endocytic compartments of RPE cells, which persisted despite attempts to clear it through upregulation of lysosomal cathepsin B, revealing a novel mechanism of lysosomal impairment in retinal degeneration. The rapid upregulation of cathepsin B was out of step with the prolonged accumulation of Aβ within lysosomes, and contrasted with enzymatic responses to internalized photoreceptor outer segments (POS). Furthermore, RPE cells exposed to Aβ were identified as deficient in cargo-carrying lysosomes at time points that are critical to POS degradation. These findings imply that Aβ accumulation within late-endocytic compartments, as well as lysosomal deficiency, impairs RPE function over time, contributing to visual defects seen in aging and AMD eyes.
Keywords: age-related macular degeneration (AMD); aging; amyloid beta (Aβ); autophagy–lysosomal pathway; retinal pigment epithelium (RPE); sight loss.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Holz F.G., Sadda S.R., Staurenghi G., Lindner M., Bird A.C., Blodi B.A., Bottoni F., Chakravarthy U., Chew E.Y., Csaky K., et al. Imaging Protocols in Clinical Studies in Advanced Age-Related Macular Degeneration: Recommendations from Classification of Atrophy Consensus Meetings. Ophthalmology. 2017;124:464–478. doi: 10.1016/j.ophtha.2016.12.002. - DOI - PubMed
-
- Holz F.G., Bindewald-Wittich A., Fleckenstein M., Dreyhaupt J., Scholl H.P., Schmitz-Valckenberg S. Progression of geographic atrophy and impact of fundus autofluorescence patterns in age-related macular degeneration. Am. J. Ophthalmol. 2007;143:463–472. doi: 10.1016/j.ajo.2006.11.041. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
