

Mechanical Mechanisms of Chromosome Segregation
- PMID: 33671543
- PMCID: PMC7926803
- DOI: 10.3390/cells10020465
Mechanical Mechanisms of Chromosome Segregation
Abstract
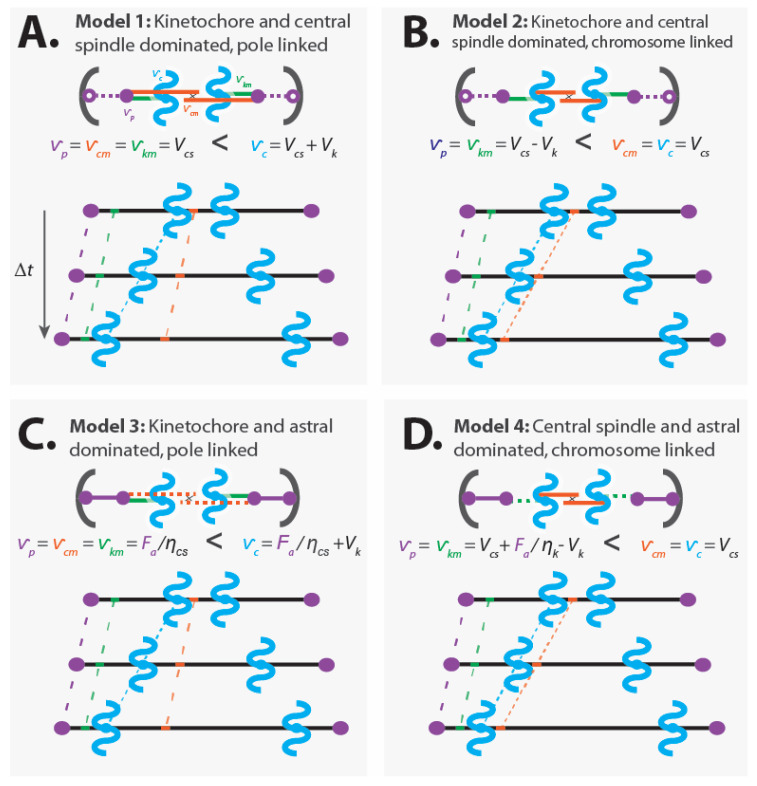
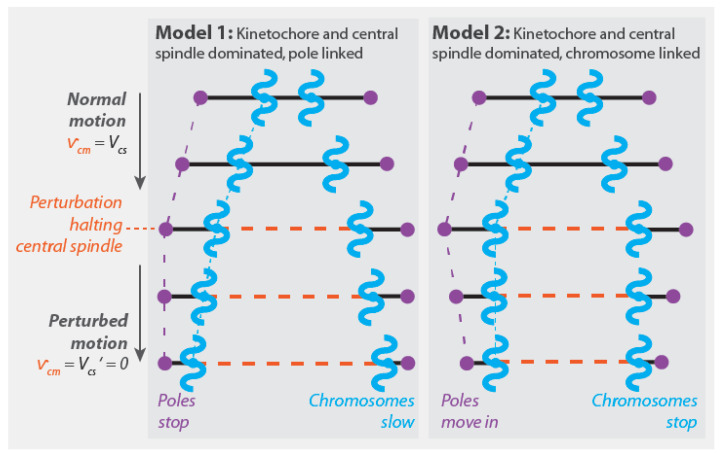
Chromosome segregation-the partitioning of genetic material into two daughter cells-is one of the most crucial processes in cell division. In all Eukaryotes, chromosome segregation is driven by the spindle, a microtubule-based, self-organizing subcellular structure. Extensive research performed over the past 150 years has identified numerous commonalities and contrasts between spindles in different systems. In this review, we use simple coarse-grained models to organize and integrate previous studies of chromosome segregation. We discuss sites of force generation in spindles and fundamental mechanical principles that any understanding of chromosome segregation must be based upon. We argue that conserved sites of force generation may interact differently in different spindles, leading to distinct mechanical mechanisms of chromosome segregation. We suggest experiments to determine which mechanical mechanism is operative in a particular spindle under study. Finally, we propose that combining biophysical experiments, coarse-grained theories, and evolutionary genetics will be a productive approach to enhance our understanding of chromosome segregation in the future.
Keywords: anaphase; chromosome segregation; mechanics; spindle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
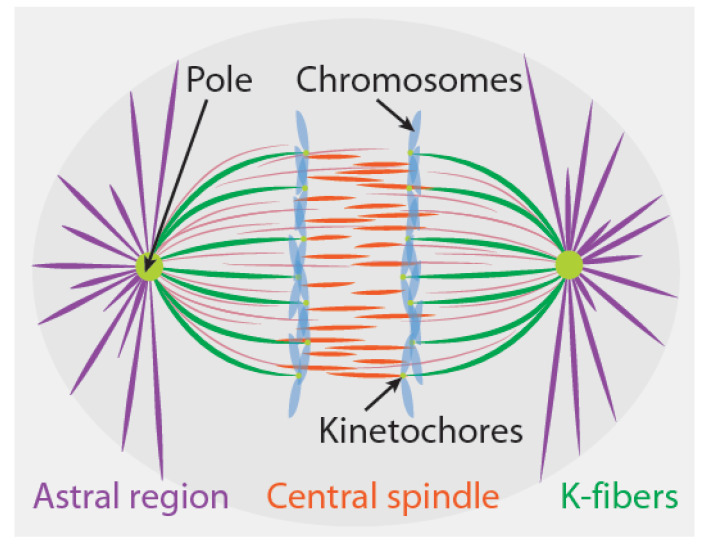
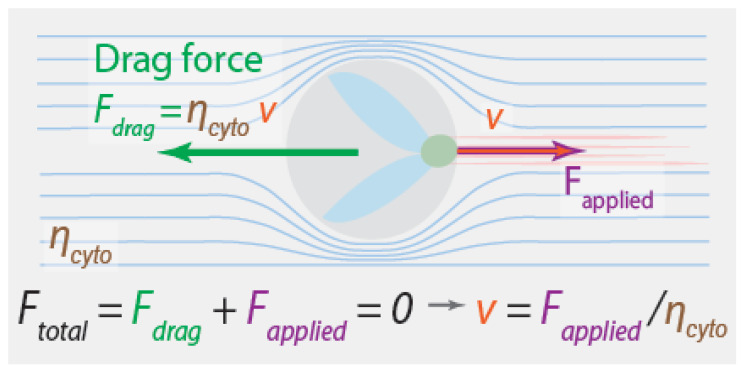
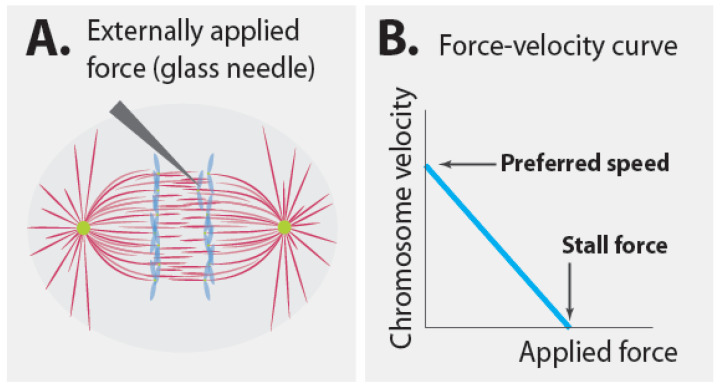
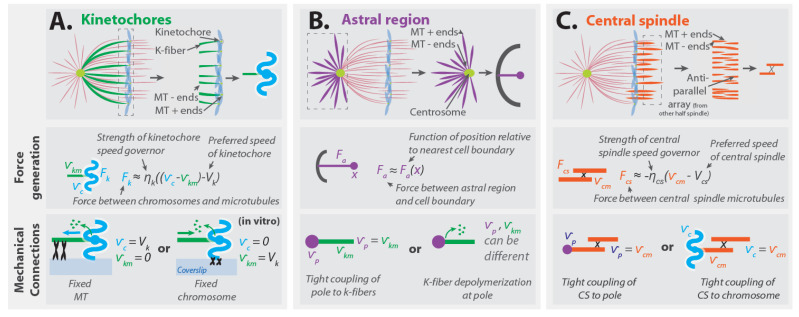
References
-
- Weinberg R.A. The Biology of Cancer. 2nd illus. ed. Garland Science; New York, NY, USA: 2013.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
