

Integrin-Linked Kinase Links Integrin Activation to Invadopodia Function and Invasion via the p(T567)-Ezrin/NHERF1/NHE1 Pathway
- PMID: 33671549
- PMCID: PMC7926356
- DOI: 10.3390/ijms22042162
Integrin-Linked Kinase Links Integrin Activation to Invadopodia Function and Invasion via the p(T567)-Ezrin/NHERF1/NHE1 Pathway
Abstract
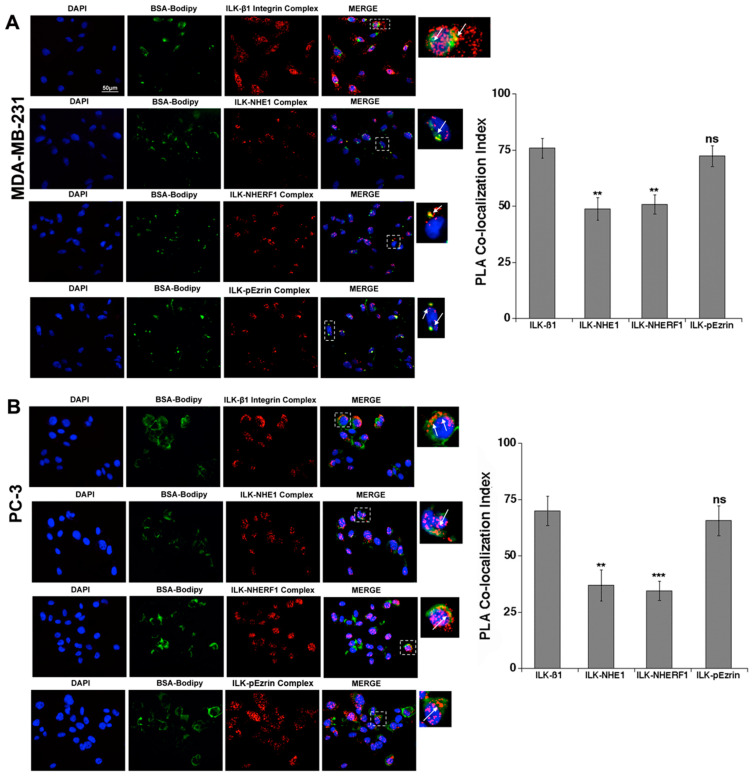
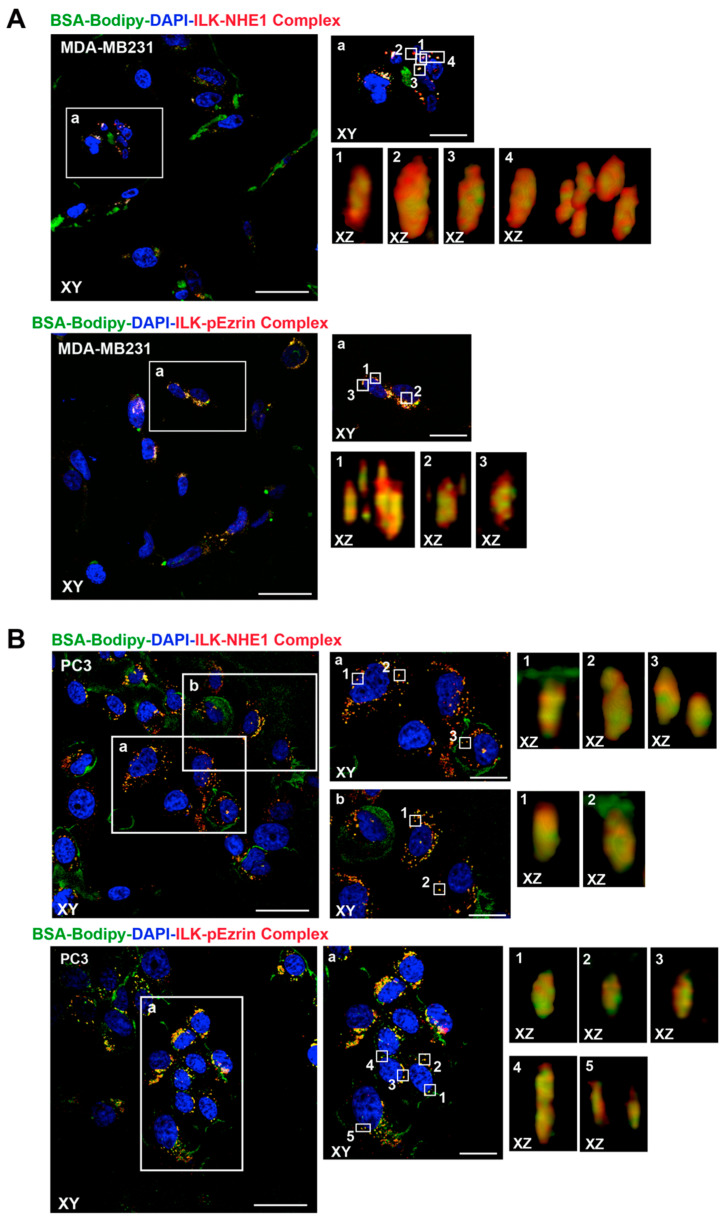
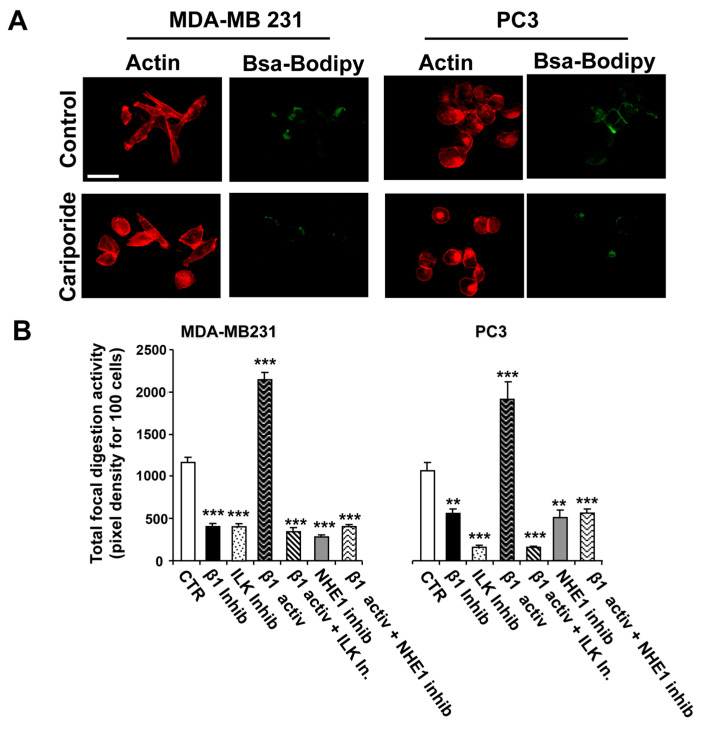
Tumor cell invasion depends largely on degradation of the extracellular matrix (ECM) by protease-rich structures called invadopodia, whose formation and activity requires the convergence of signaling pathways engaged in cell adhesion, actin assembly, membrane regulation and ECM proteolysis. It is known that β1-integrin stimulates invadopodia function through an invadopodial p(T567)-ezrin/NHERF1/NHE1 signal complex that regulates NHE1-driven invadopodia proteolytic activity and invasion. However, the link between β1-integrin and this signaling complex is unknown. In this study, in metastatic breast (MDA-MB-231) and prostate (PC-3) cancer cells, we report that integrin-linked kinase (ILK) integrates β1-integrin with this signaling complex to regulate invadopodia activity and invasion. Proximity ligation assay experiments demonstrate that, in invadopodia, ILK associates with β1-integrin, NHE1 and the scaffold proteins p(T567)-ezrin and NHERF1. Activation of β1-integrin increased both invasion and invadopodia activity, which were specifically blocked by inhibition of either NHE1 or ILK. We conclude that ILK integrates β1-integrin with the ECM proteolytic/invasion signal module to induce NHE1-driven invadopodial ECM proteolysis and cell invasion.
Keywords: breast cancer; integrin signaling; invadopodia; invasion; pH; prostate cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
