

Divalent Cation Modulation of Ion Permeation in TMEM16 Proteins
- PMID: 33672260
- PMCID: PMC7926781
- DOI: 10.3390/ijms22042209
Divalent Cation Modulation of Ion Permeation in TMEM16 Proteins
Abstract
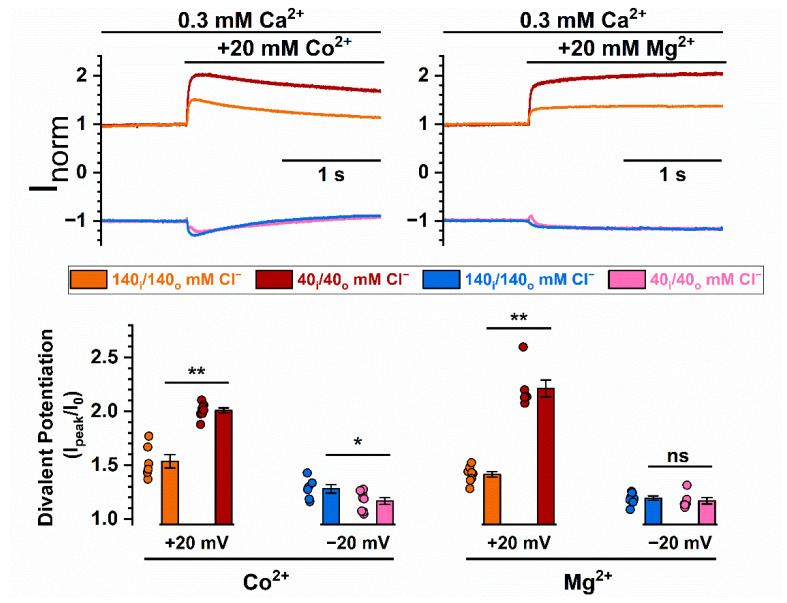
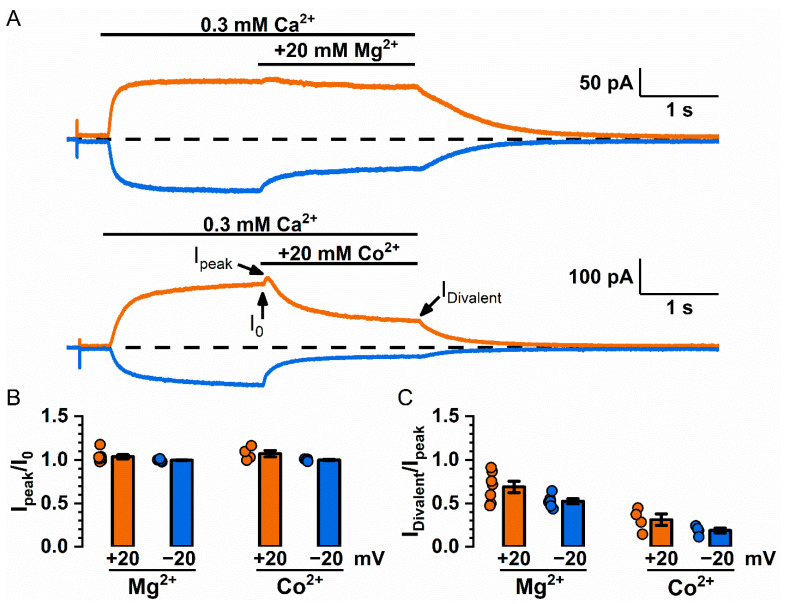
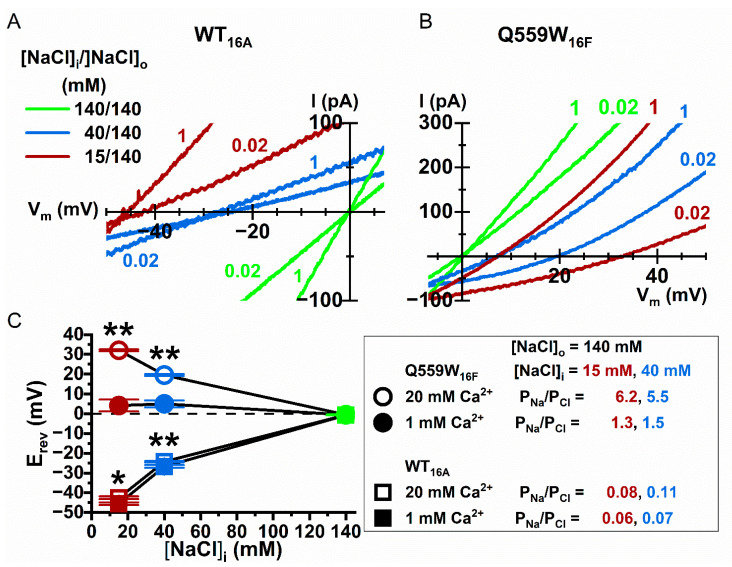
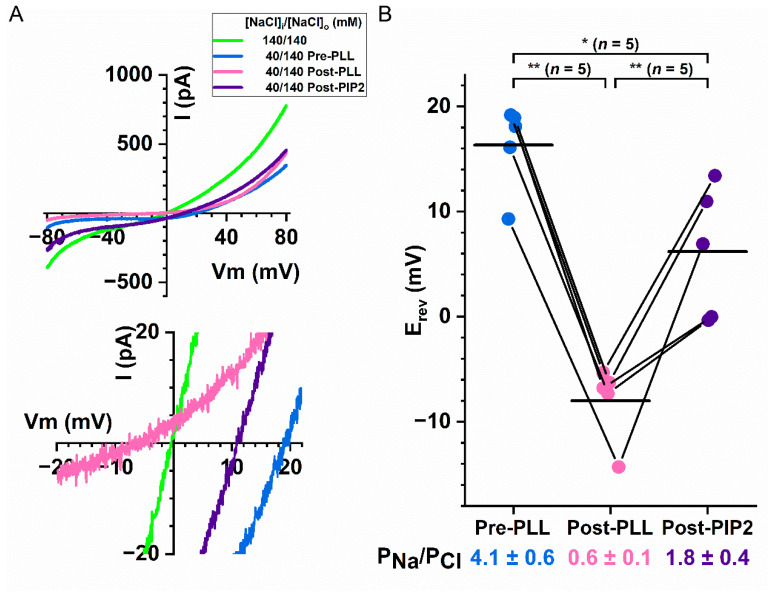
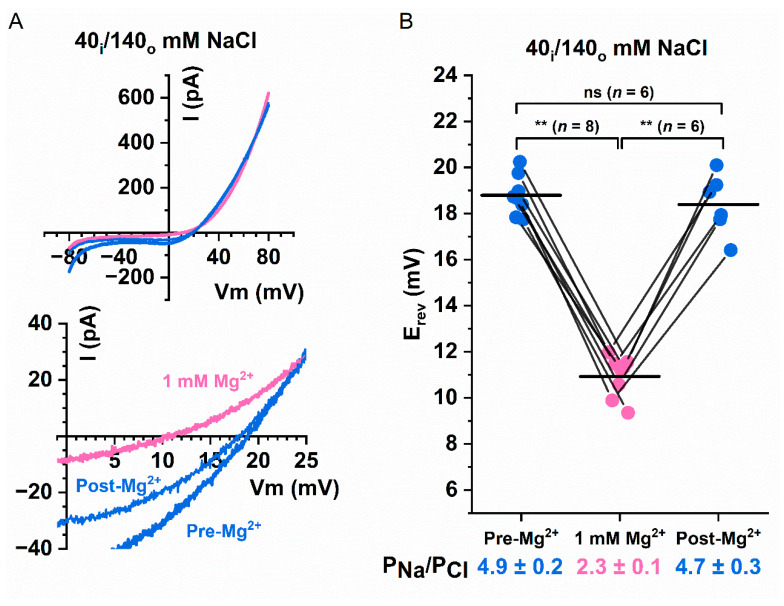
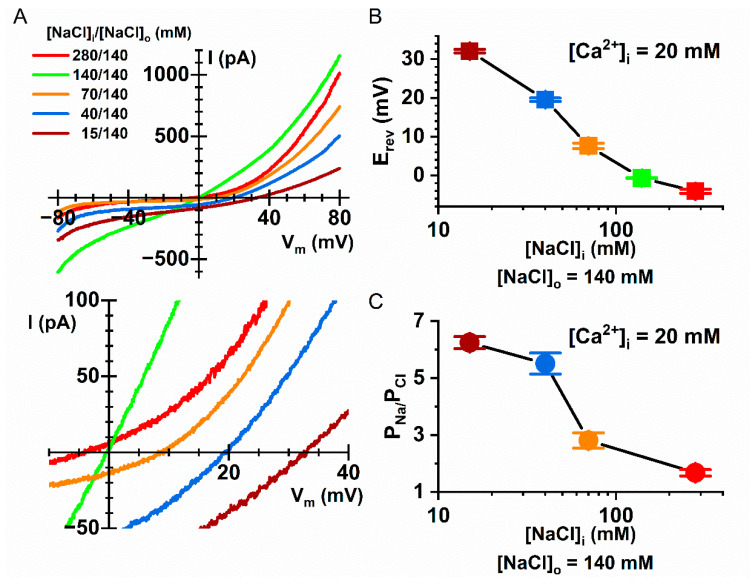
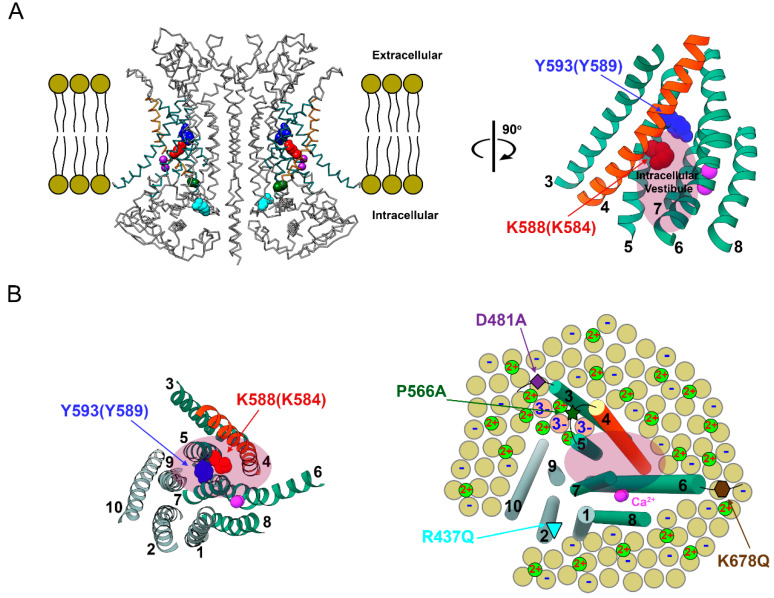
Intracellular divalent cations control the molecular function of transmembrane protein 16 (TMEM16) family members. Both anion channels (such as TMEM16A) and phospholipid scramblases (such as TMEM16F) in this family are activated by intracellular Ca2+ in the low µM range. In addition, intracellular Ca2+ or Co2+ at mM concentrations have been shown to further potentiate the saturated Ca2+-activated current of TMEM16A. In this study, we found that all alkaline earth divalent cations in mM concentrations can generate similar potentiation effects in TMEM16A when applied intracellularly, and that manipulations thought to deplete membrane phospholipids weaken the effect. In comparison, mM concentrations of divalent cations minimally potentiate the current of TMEM16F but significantly change its cation/anion selectivity. We suggest that divalent cations may increase local concentrations of permeant ions via a change in pore electrostatic potential, possibly acting through phospholipid head groups in or near the pore. Monovalent cations appear to exert a similar effect, although with a much lower affinity. Our findings resolve controversies regarding the ion selectivity of TMEM16 proteins. The physiological role of this mechanism, however, remains elusive because of the nearly constant high cation concentrations in cytosols.
Keywords: TMEM16A; TMEM16F; divalent cations; permeability ratio; phospholipids.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
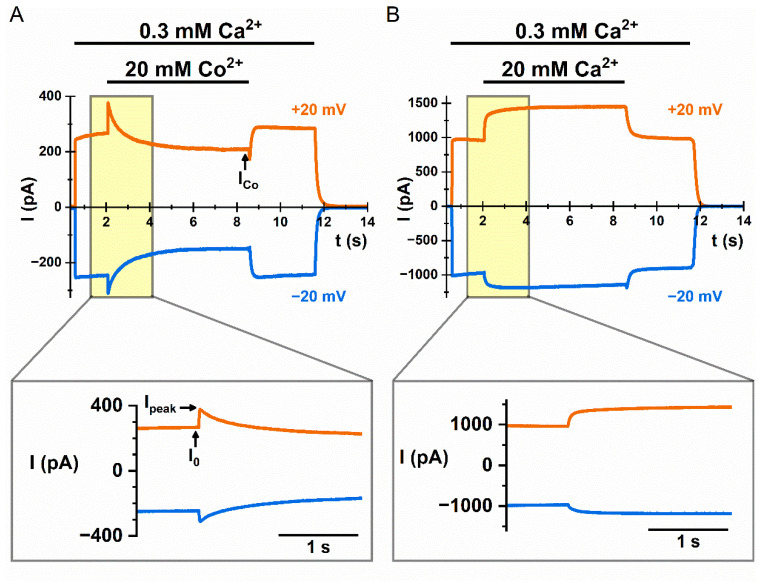
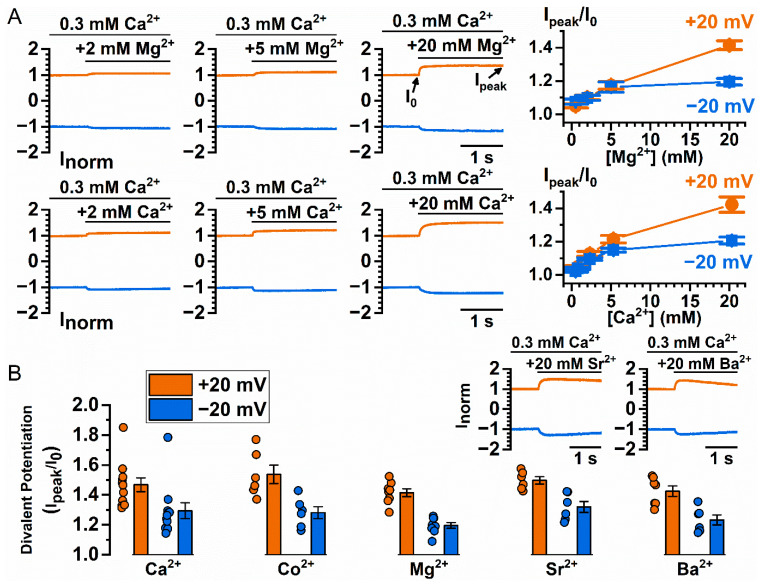
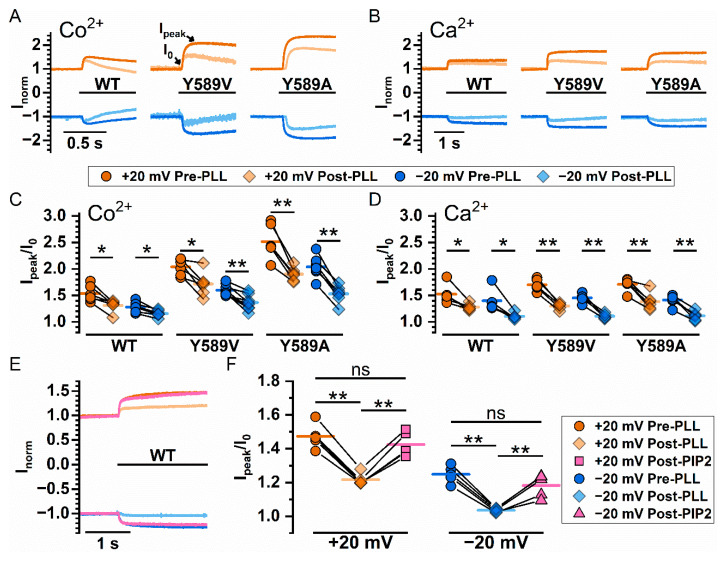
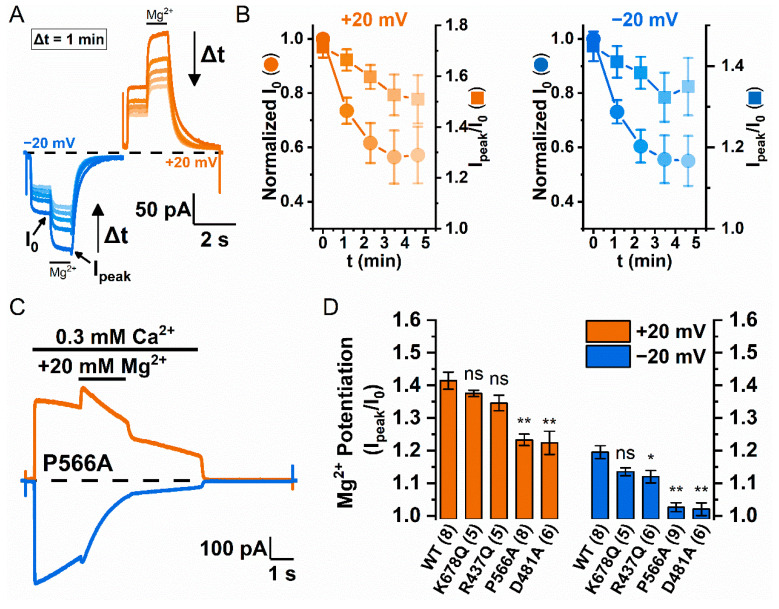
Similar articles
-
Structure and Function of Calcium-Activated Chloride Channels and Phospholipid Scramblases in the TMEM16 Family.Handb Exp Pharmacol. 2024;283:153-180. doi: 10.1007/164_2022_595. Handb Exp Pharmacol. 2024. PMID: 35792944 Review.
-
Regulation of TMEM16A/ANO1 and TMEM16F/ANO6 ion currents and phospholipid scrambling by Ca2+ and plasma membrane lipid.J Physiol. 2018 Jan 15;596(2):217-229. doi: 10.1113/JP275175. Epub 2017 Dec 18. J Physiol. 2018. PMID: 29134661 Free PMC article.
-
Cobalt ion interaction with TMEM16A calcium-activated chloride channel: Inhibition and potentiation.PLoS One. 2020 Apr 17;15(4):e0231812. doi: 10.1371/journal.pone.0231812. eCollection 2020. PLoS One. 2020. PMID: 32302365 Free PMC article.
-
Characterization of the scrambling domain of the TMEM16 family.Proc Natl Acad Sci U S A. 2017 Jun 13;114(24):6274-6279. doi: 10.1073/pnas.1703391114. Epub 2017 May 30. Proc Natl Acad Sci U S A. 2017. PMID: 28559311 Free PMC article.
-
Anoctamin pharmacology.Cell Calcium. 2024 Jul;121:102905. doi: 10.1016/j.ceca.2024.102905. Epub 2024 May 10. Cell Calcium. 2024. PMID: 38788257 Review.
Cited by
-
Chloride ions in health and disease.Biosci Rep. 2024 May 29;44(5):BSR20240029. doi: 10.1042/BSR20240029. Biosci Rep. 2024. PMID: 38573803 Free PMC article. Review.
-
Structural heterogeneity of the ion and lipid channel TMEM16F.Nat Commun. 2024 Jan 2;15(1):110. doi: 10.1038/s41467-023-44377-7. Nat Commun. 2024. PMID: 38167485 Free PMC article.
-
Structure and Function of Calcium-Activated Chloride Channels and Phospholipid Scramblases in the TMEM16 Family.Handb Exp Pharmacol. 2024;283:153-180. doi: 10.1007/164_2022_595. Handb Exp Pharmacol. 2024. PMID: 35792944 Review.
-
Roles, molecular mechanisms, and signaling pathways of TMEMs in neurological diseases.Am J Transl Res. 2021 Dec 15;13(12):13273-13297. eCollection 2021. Am J Transl Res. 2021. PMID: 35035675 Free PMC article. Review.
-
Simulation-based survey of TMEM16 family reveals that robust lipid scrambling requires an open groove.bioRxiv [Preprint]. 2025 Mar 29:2024.09.25.615027. doi: 10.1101/2024.09.25.615027. bioRxiv. 2025. Update in: Elife. 2025 Aug 06;14:RP105111. doi: 10.7554/eLife.105111. PMID: 39386458 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
