

Recent Advances in Bunyavirus Glycoprotein Research: Precursor Processing, Receptor Binding and Structure
- PMID: 33672327
- PMCID: PMC7926653
- DOI: 10.3390/v13020353
Recent Advances in Bunyavirus Glycoprotein Research: Precursor Processing, Receptor Binding and Structure
Abstract
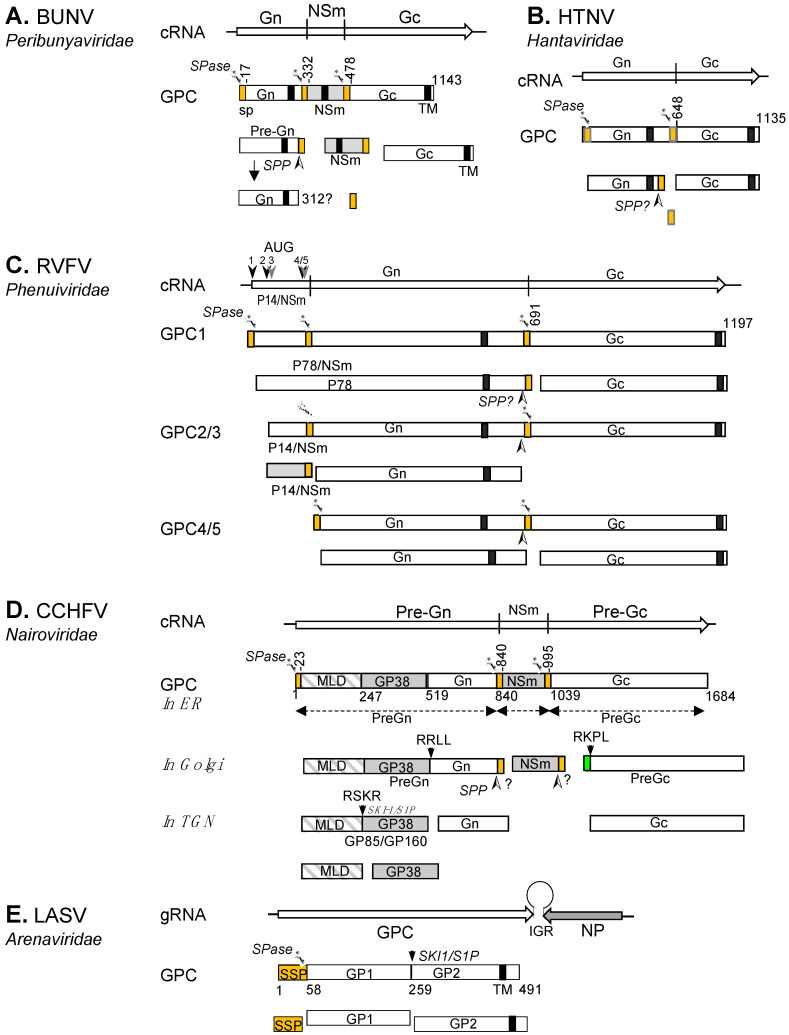
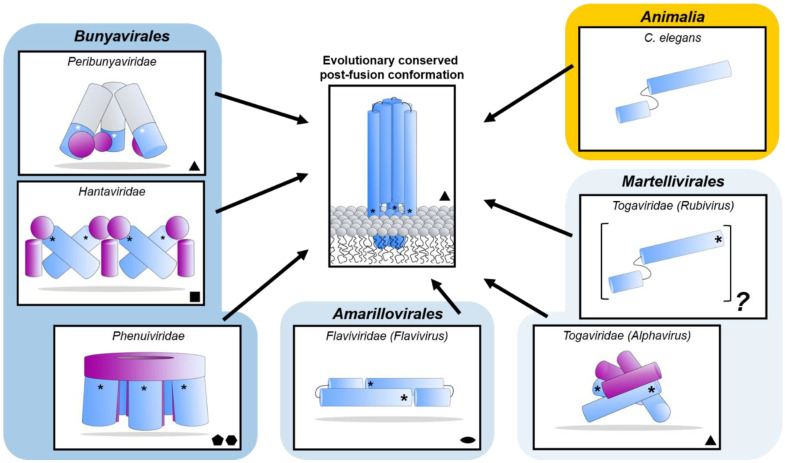
The Bunyavirales order accommodates related viruses (bunyaviruses) with segmented, linear, single-stranded, negative- or ambi-sense RNA genomes. Their glycoproteins form capsomeric projections or spikes on the virion surface and play a crucial role in virus entry, assembly, morphogenesis. Bunyavirus glycoproteins are encoded by a single RNA segment as a polyprotein precursor that is co- and post-translationally cleaved by host cell enzymes to yield two mature glycoproteins, Gn and Gc (or GP1 and GP2 in arenaviruses). These glycoproteins undergo extensive N-linked glycosylation and despite their cleavage, remain associated to the virion to form an integral transmembrane glycoprotein complex. This review summarizes recent advances in our understanding of the molecular biology of bunyavirus glycoproteins, including their processing, structure, and known interactions with host factors that facilitate cell entry.
Keywords: Bunyavirales; bunyavirus; genomic coding strategy; glycoprotein precursor processing; glycoproteins; protein structure; receptor binding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The Postfusion Structure of the Heartland Virus Gc Glycoprotein Supports Taxonomic Separation of the Bunyaviral Families Phenuiviridae and Hantaviridae.J Virol. 2017 Dec 14;92(1):e01558-17. doi: 10.1128/JVI.01558-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29070692 Free PMC article.
-
Mechanisms of bunyavirus morphogenesis and egress.J Gen Virol. 2023 Apr;104(4). doi: 10.1099/jgv.0.001845. J Gen Virol. 2023. PMID: 37083579 Review.
-
Mutations in the Schmallenberg Virus Gc Glycoprotein Facilitate Cellular Protein Synthesis Shutoff and Restore Pathogenicity of NSs Deletion Mutants in Mice.J Virol. 2016 May 12;90(11):5440-5450. doi: 10.1128/JVI.00424-16. Print 2016 Jun 1. J Virol. 2016. PMID: 26984728 Free PMC article.
-
Analysis of the humoral immune response against the envelope glycoprotein Gc of Schmallenberg virus reveals a domain located at the amino terminus targeted by mAbs with neutralizing activity.J Gen Virol. 2016 Mar;97(3):571-580. doi: 10.1099/jgv.0.000377. Epub 2015 Dec 18. J Gen Virol. 2016. PMID: 26684324
-
The Envelope Proteins of the Bunyavirales.Adv Virus Res. 2017;98:83-118. doi: 10.1016/bs.aivir.2017.02.002. Epub 2017 Apr 8. Adv Virus Res. 2017. PMID: 28433053 Review.
Cited by
-
Discovery and Genomic Function of a Novel Rice Dwarf-Associated Bunya-like Virus.Viruses. 2022 May 29;14(6):1183. doi: 10.3390/v14061183. Viruses. 2022. PMID: 35746655 Free PMC article.
-
Structural basis of synergistic neutralization of Crimean-Congo hemorrhagic fever virus by human antibodies.Science. 2022 Jan 7;375(6576):104-109. doi: 10.1126/science.abl6502. Epub 2021 Nov 18. Science. 2022. PMID: 34793197 Free PMC article.
-
Structure and stabilization of the antigenic glycoprotein building blocks of the New World mammarenavirus spike complex.mBio. 2025 Jul 9;16(7):e0107625. doi: 10.1128/mbio.01076-25. Epub 2025 Jun 13. mBio. 2025. PMID: 40511941 Free PMC article.
-
Potent immunogenicity and neutralization of recombinant adeno-associated virus expressing the glycoprotein of severe fever with thrombocytopenia virus.J Vet Med Sci. 2024 Feb 15;86(2):228-238. doi: 10.1292/jvms.23-0375. Epub 2023 Dec 22. J Vet Med Sci. 2024. PMID: 38143087 Free PMC article.
-
NMR side-chain assignments of the Crimean-Congo hemorrhagic fever virus glycoprotein n cytosolic domain.Magn Reson (Gott). 2024 Jul 8;5(2):95-101. doi: 10.5194/mr-5-95-2024. eCollection 2024. Magn Reson (Gott). 2024. PMID: 40521231 Free PMC article.
References
-
- Kuhn J.H., Adkins S., Alioto D., Alkhovsky S.V., Amarasinghe G.K., Anthony S.J., Avšič-Županc T., Ayllón M.A., Bahl J., Balkema-Buschmann A., et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020;165:3023–3072. doi: 10.1007/s00705-020-04731-2. - DOI - PMC - PubMed
-
- Plyusnin A., Beaty B.J., Elliott R.M., Kormelink R., Lundkvist A., Schmaljohn C.S., Tesh R.B. Bunyaviridae. In: Andrew M.Q., King M.J.A., Eric B.C., Lefkowitz E.J., editors. Virus Taxonomy. Elsevier; Oxford, UK: 2011. pp. 725–742.
-
- Elliott R.M., Schmaljohn C.S. Bunyaviridae. In: Knipe D.M., Howley P.M., editors. Fields Virology. 6th ed. Wolters Kluwer; Philadelphia, PA, USA: 2013. pp. 1244–1282.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
