

Valency and Binding Affinity Variations Can Regulate the Multilayered Organization of Protein Condensates with Many Components
- PMID: 33672806
- PMCID: PMC7918469
- DOI: 10.3390/biom11020278
Valency and Binding Affinity Variations Can Regulate the Multilayered Organization of Protein Condensates with Many Components
Abstract
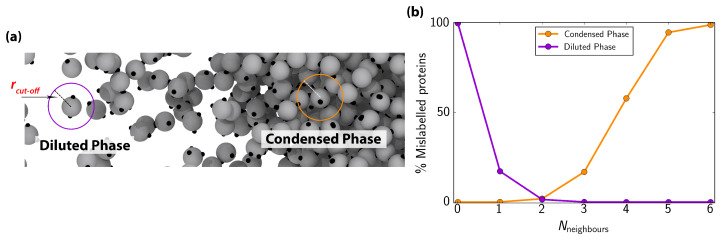
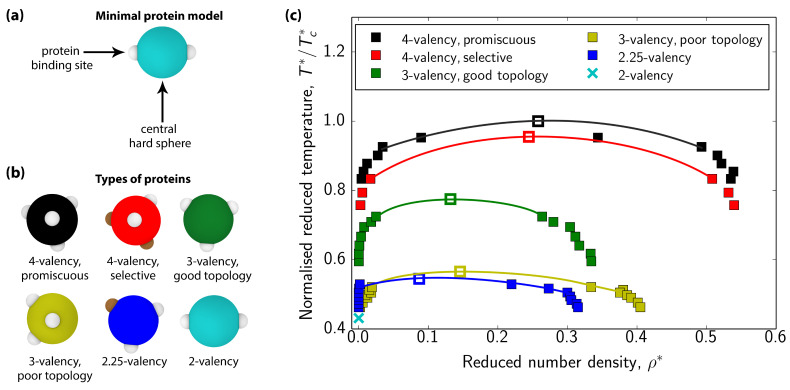
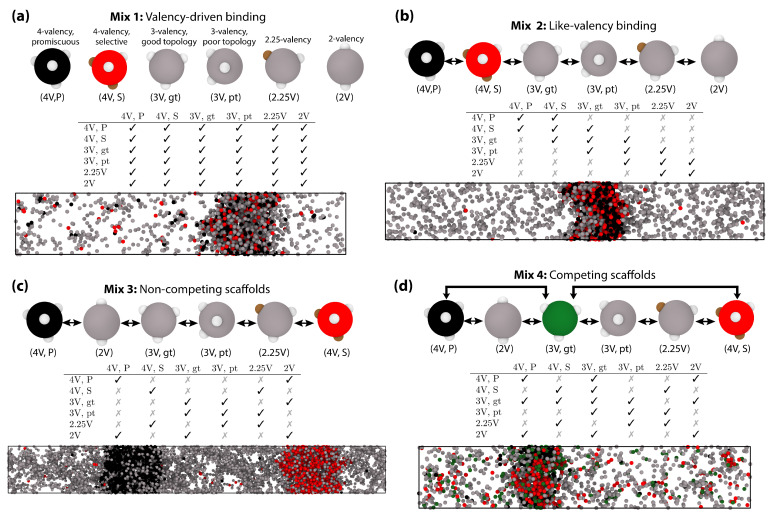
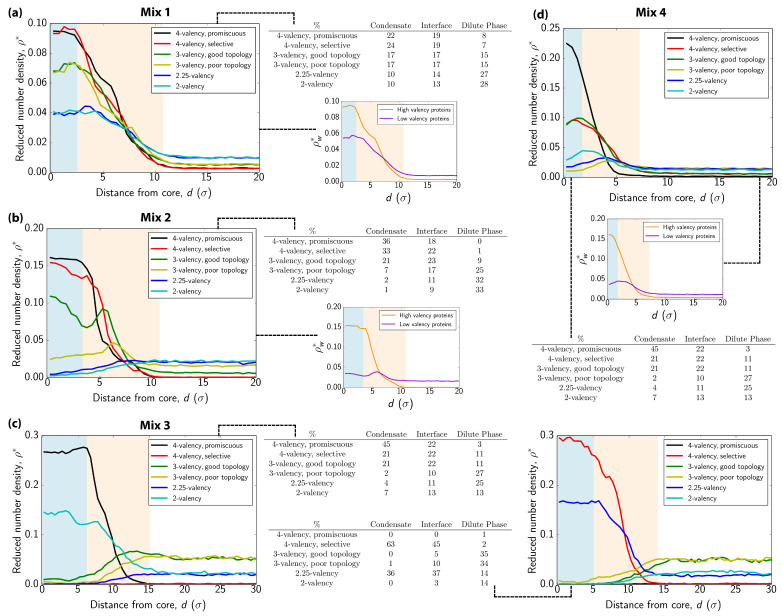
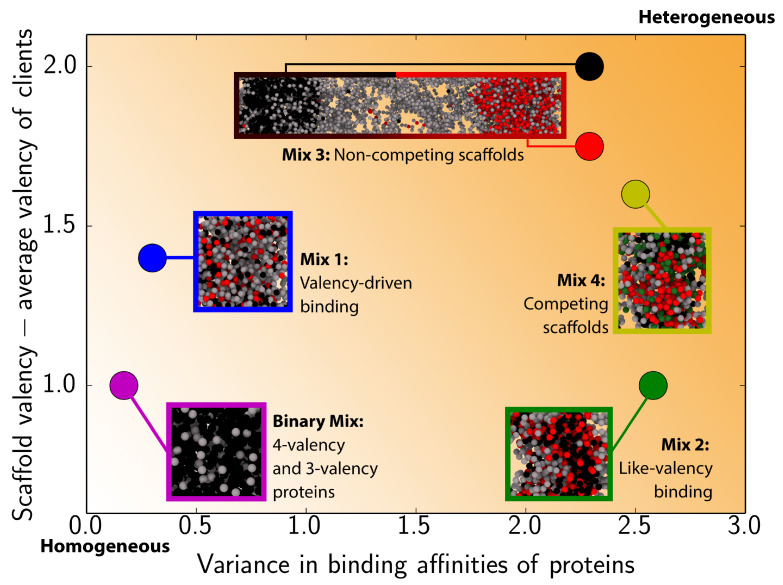
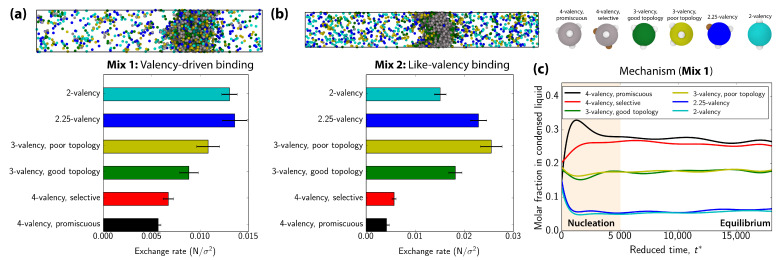
Biomolecular condensates, which assemble via the process of liquid-liquid phase separation (LLPS), are multicomponent compartments found ubiquitously inside cells. Experiments and simulations have shown that biomolecular condensates with many components can exhibit multilayered organizations. Using a minimal coarse-grained model for interacting multivalent proteins, we investigate the thermodynamic parameters governing the formation of multilayered condensates through changes in protein valency and binding affinity. We focus on multicomponent condensates formed by scaffold proteins (high-valency proteins that can phase separate on their own via homotypic interactions) and clients (proteins recruited to condensates via heterotypic scaffold-client interactions). We demonstrate that higher valency species are sequestered to the center of the multicomponent condensates, while lower valency proteins cluster towards the condensate interface. Such multilayered condensate architecture maximizes the density of LLPS-stabilizing molecular interactions, while simultaneously reducing the surface tension of the condensates. In addition, multilayered condensates exhibit rapid exchanges of low valency proteins in and out, while keeping higher valency proteins-the key biomolecules involved in condensate nucleation-mostly within. We also demonstrate how modulating the binding affinities among the different proteins in a multicomponent condensate can significantly transform its multilayered structure, and even trigger fission of a condensate into multiple droplets with different compositions.
Keywords: minimal protein model; multicomponent condensates; multilayered condensates; multiphase condensates; protein liquid–liquid phase separation.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Sear R.P. The cytoplasm of living cells: A functional mixture of thousands of components. J. Phys. Condens. Matter. 2005;17:S3587–S3595. doi: 10.1088/0953-8984/17/45/052. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
