

Intestinal Epithelial Barrier Maturation by Enteric Glial Cells Is GDNF-Dependent
- PMID: 33672854
- PMCID: PMC7917776
- DOI: 10.3390/ijms22041887
Intestinal Epithelial Barrier Maturation by Enteric Glial Cells Is GDNF-Dependent
Abstract
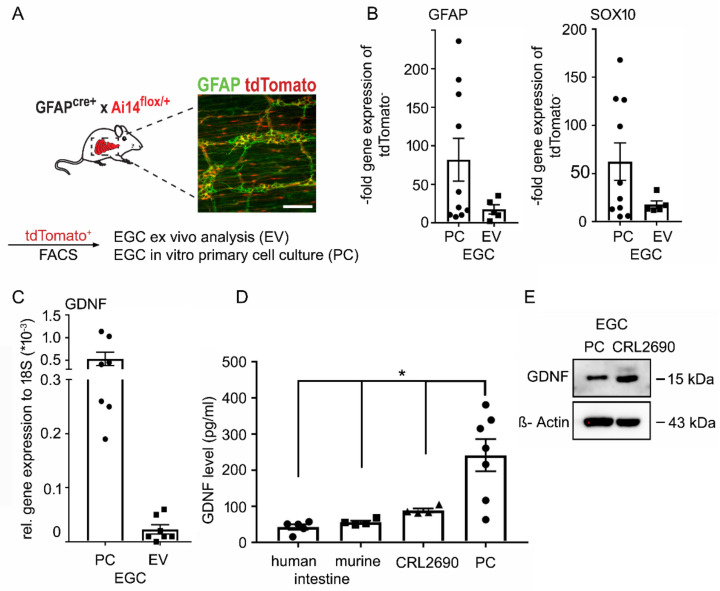
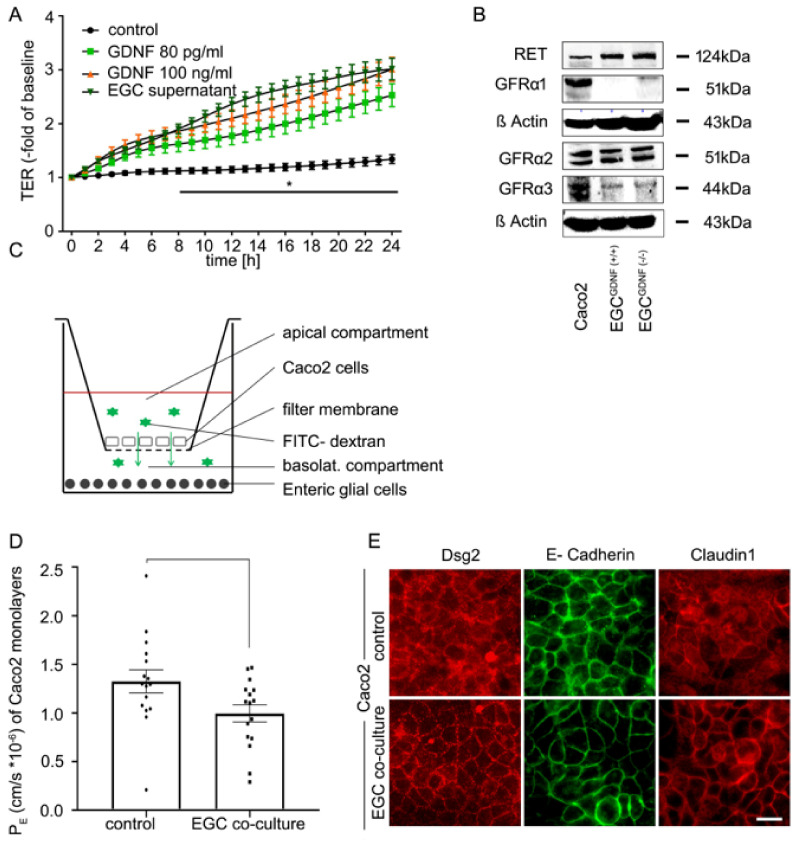
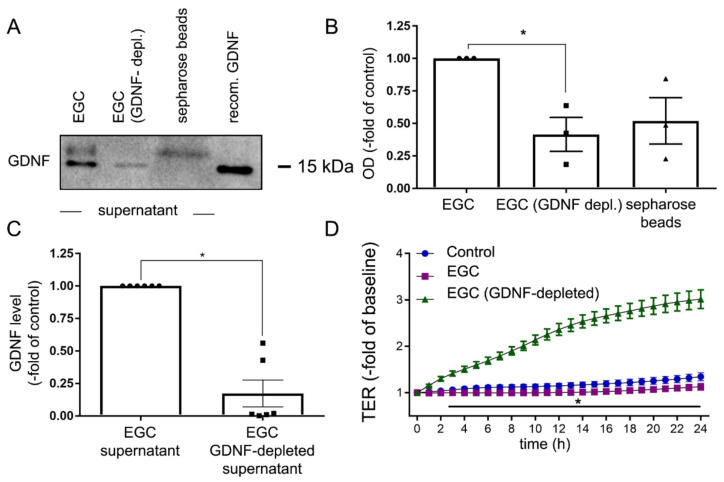
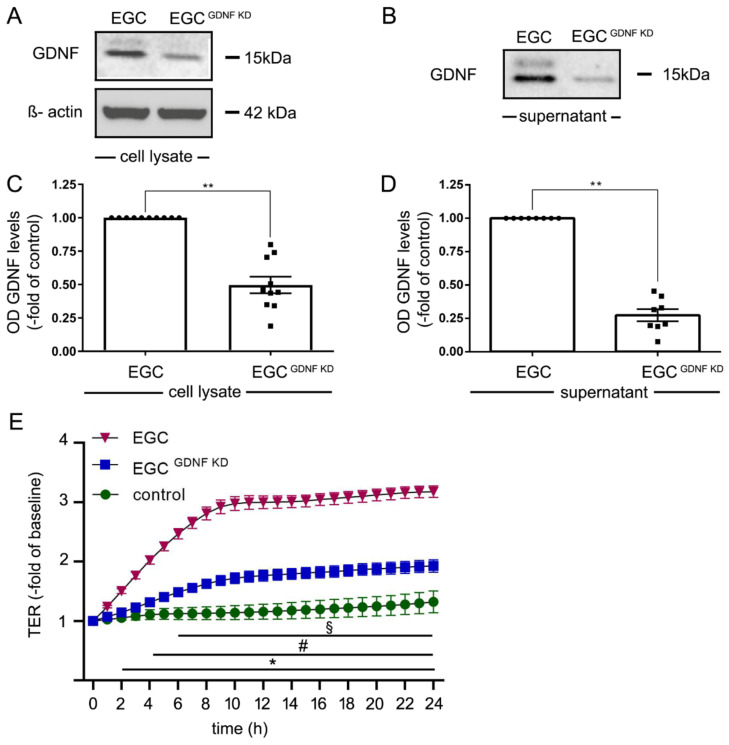
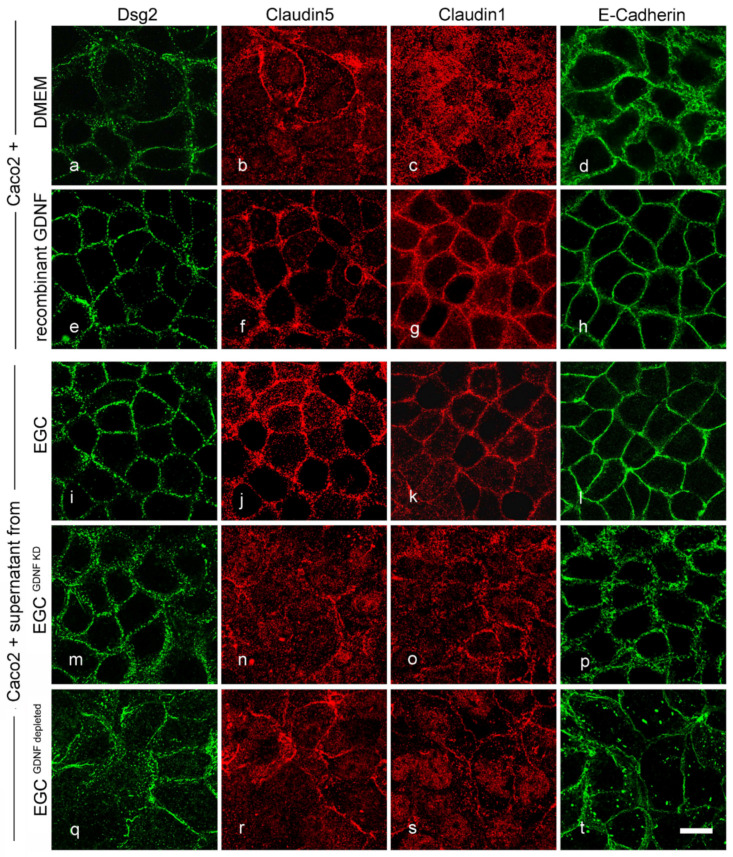
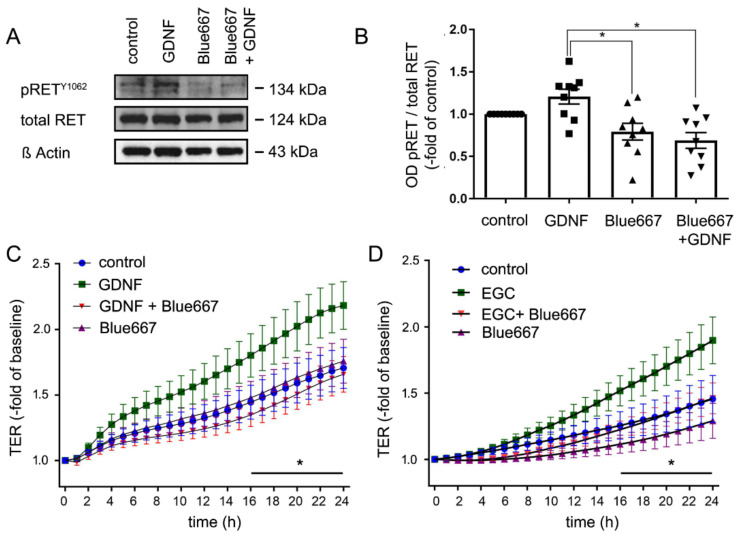
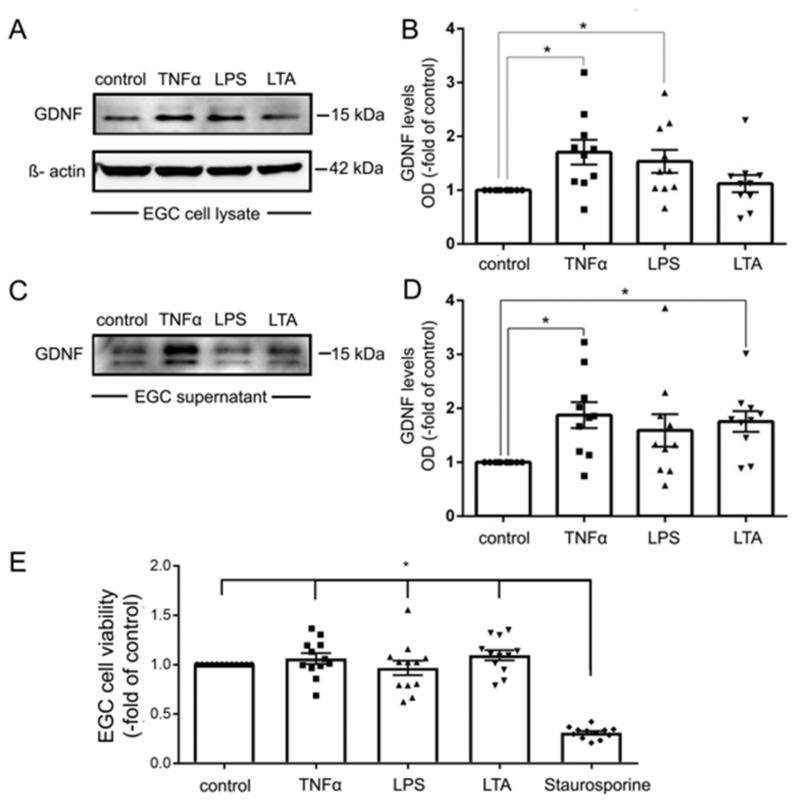
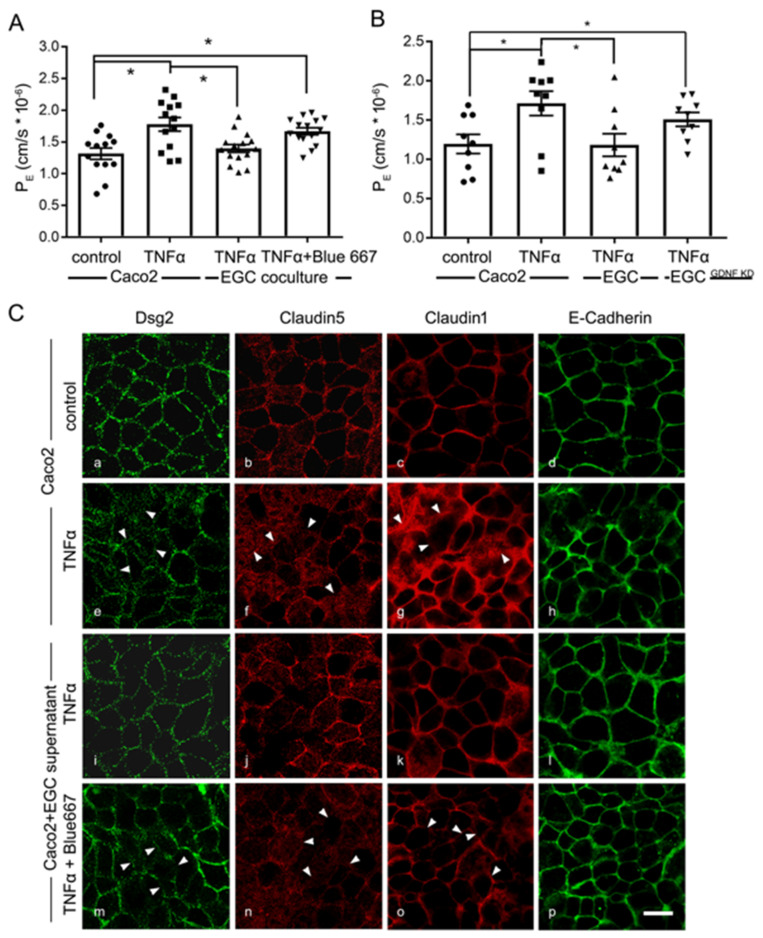
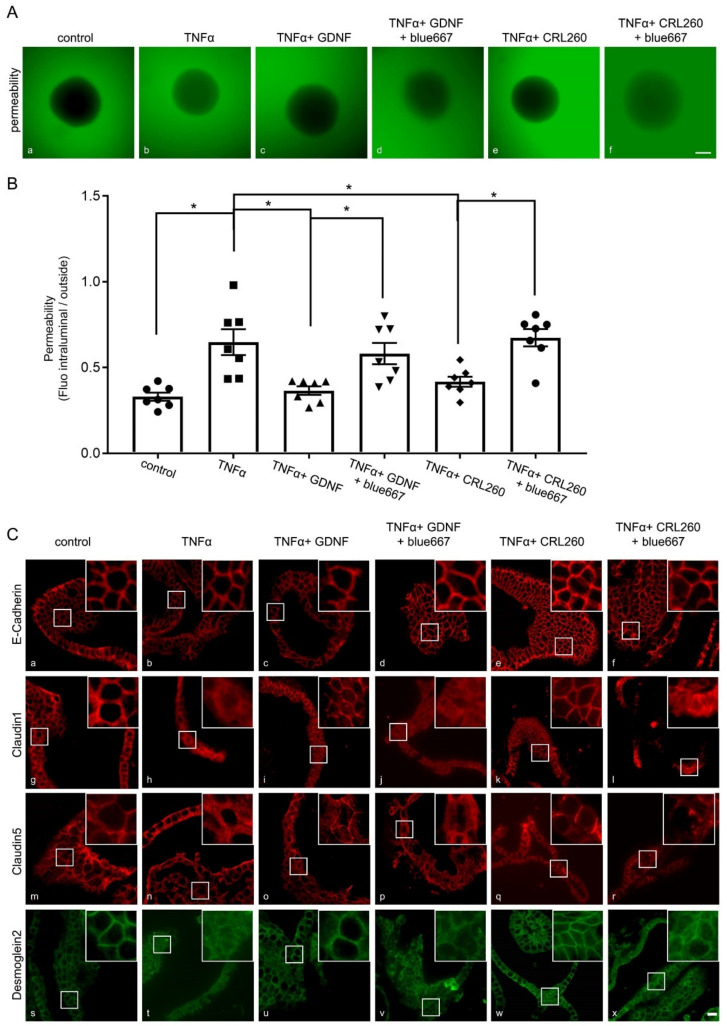
Enteric glial cells (EGCs) of the enteric nervous system are critically involved in the maintenance of intestinal epithelial barrier function (IEB). The underlying mechanisms remain undefined. Glial cell line-derived neurotrophic factor (GDNF) contributes to IEB maturation and may therefore be the predominant mediator of this process by EGCs. Using GFAPcre x Ai14floxed mice to isolate EGCs by Fluorescence-activated cell sorting (FACS), we confirmed that they synthesize GDNF in vivo as well as in primary cultures demonstrating that EGCs are a rich source of GDNF in vivo and in vitro. Co-culture of EGCs with Caco2 cells resulted in IEB maturation which was abrogated when GDNF was either depleted from EGC supernatants, or knocked down in EGCs or when the GDNF receptor RET was blocked. Further, TNFα-induced loss of IEB function in Caco2 cells and in organoids was attenuated by EGC supernatants or by recombinant GDNF. These barrier-protective effects were blunted when using supernatants from GDNF-deficient EGCs or by RET receptor blockade. Together, our data show that EGCs produce GDNF to maintain IEB function in vitro through the RET receptor.
Keywords: GDNF5; RET6; enteric glial cells; enteric nervous system; gut barrier; inflammatory bowel disease; intercellular junctions; intestinal epithelial barrier; neurotrophic factors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
GDNF is involved in the barrier-inducing effect of enteric glial cells on intestinal epithelial cells under acute ischemia reperfusion stimulation.Mol Neurobiol. 2014 Oct;50(2):274-89. doi: 10.1007/s12035-014-8730-9. Epub 2014 May 31. Mol Neurobiol. 2014. PMID: 24878766
-
Enteric glial-mediated enhancement of intestinal barrier integrity is compromised by morphine.J Surg Res. 2017 Nov;219:214-221. doi: 10.1016/j.jss.2017.05.099. Epub 2017 Jun 30. J Surg Res. 2017. PMID: 29078884 Free PMC article.
-
Defects in 15-HETE Production and Control of Epithelial Permeability by Human Enteric Glial Cells From Patients With Crohn's Disease.Gastroenterology. 2016 Jan;150(1):168-80. doi: 10.1053/j.gastro.2015.09.038. Epub 2015 Nov 11. Gastroenterology. 2016. PMID: 26433161
-
The Involvement of Glial Cell-Derived Neurotrophic Factor in Inflammatory Bowel Disease.J Interferon Cytokine Res. 2022 Jan;42(1):1-7. doi: 10.1089/jir.2021.0116. Epub 2021 Nov 26. J Interferon Cytokine Res. 2022. PMID: 34846920 Review.
-
The glial cell-line derived neurotrophic factor: a novel regulator of intestinal barrier function in health and disease.Am J Physiol Gastrointest Liver Physiol. 2016 Jun 1;310(11):G1118-23. doi: 10.1152/ajpgi.00125.2016. Epub 2016 May 5. Am J Physiol Gastrointest Liver Physiol. 2016. PMID: 27151942 Review.
Cited by
-
β-adrenergic signaling triggers enteric glial reactivity and acute enteric gliosis during surgery.J Neuroinflammation. 2023 Nov 8;20(1):255. doi: 10.1186/s12974-023-02937-0. J Neuroinflammation. 2023. PMID: 37941007 Free PMC article.
-
Organoids in gastrointestinal diseases: from bench to clinic.MedComm (2020). 2024 Jun 29;5(7):e574. doi: 10.1002/mco2.574. eCollection 2024 Jul. MedComm (2020). 2024. PMID: 38948115 Free PMC article. Review.
-
Guardians of the gut: influence of the enteric nervous system on the intestinal epithelial barrier.Front Med (Lausanne). 2023 Aug 25;10:1228938. doi: 10.3389/fmed.2023.1228938. eCollection 2023. Front Med (Lausanne). 2023. PMID: 37692784 Free PMC article. Review.
-
Glial cell line derived neurotrophic factor (GDNF) induces mucosal healing via intestinal stem cell niche activation.Cell Prolif. 2025 Feb;58(2):e13758. doi: 10.1111/cpr.13758. Epub 2024 Nov 28. Cell Prolif. 2025. PMID: 39610047 Free PMC article.
-
Aging, brain-derived neurotrophic factor, and allergen-induced pulmonary responses in mice.Am J Physiol Lung Cell Mol Physiol. 2025 Feb 1;328(2):L290-L300. doi: 10.1152/ajplung.00145.2024. Epub 2024 Oct 22. Am J Physiol Lung Cell Mol Physiol. 2025. PMID: 39437757 Free PMC article.
References
-
- Neunlist M., Toumi F., Oreschkova T., Denis M., Leborgne J., Laboisse C.L., Galmiche J.P., Jarry A. Human ENS regulates the intestinal epithelial barrier permeability and a tight junction-associated protein ZO-1 via VIPergic pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2003;285:G1028–G1036. doi: 10.1152/ajpgi.00066.2003. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
